Make more informed purchasing decisions with our new product availability and delivery estimate feature, now available on all product pages, in your cart, and during checkout.
Sign In
New to STEMCELL?
Register for an account to quickly and easily purchase products online and for one-click access to all educational content.
New look, same high quality and support! You may notice that your instrument or reagent packaging looks slightly different from images displayed on the website, or from previous orders. We are updating our look but rest assured, the products themselves and how you should use them have not changed. Learn more
Request Pricing
Thank you for your interest in this product.
Please provide us with your contact information and your local representative
will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration
By submitting this form, you are providing your consent to STEMCELL Technologies Canada Inc. and its subsidiaries and affiliates (“STEMCELL”) to collect and use your information, and send you newsletters and emails in accordance with our privacy policy. Please contact us with any questions that you may have. You can unsubscribe or change your email preferences at any time.
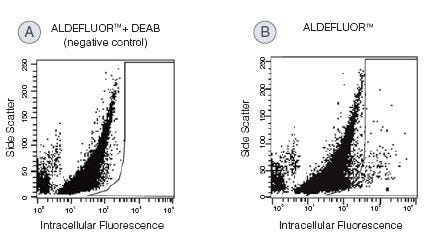
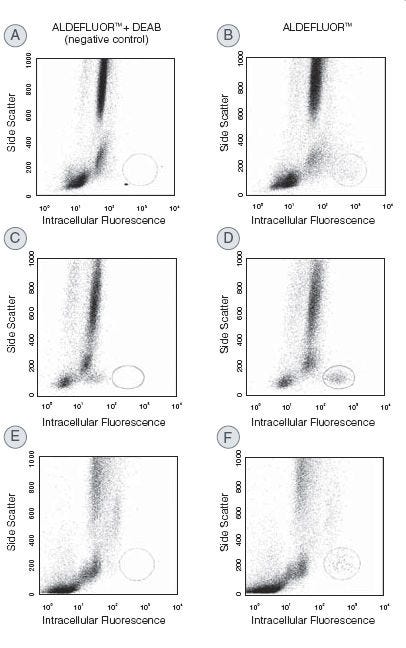
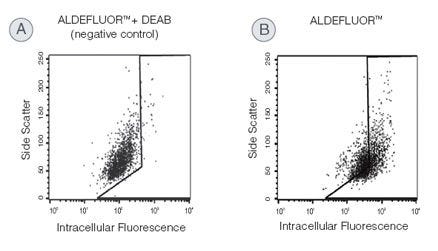
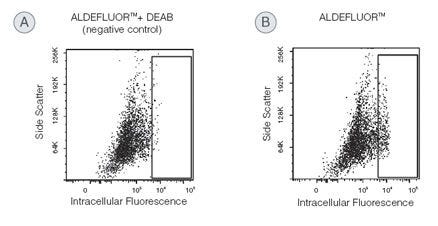
Identify and isolate viable cells expressing aldehyde dehydrogenase (ALDH) with the ALDEFLUOR™ Kit. Compared to traditional methods, this assay does not require antibody staining.
High ALDH expression has been reported for normal and cancer precursor cells of various lineages. The ALDEFLUOR™ assay is a widely published non-immunological method for the detection of ALDH-bright (ALDHbr) cells and can be used to detect cancer cells in hematopoietic, mammary, endothelial, mesenchymal, neural, and other tissues.
The kit is compatible with standard flow cytometers for performing downstream analysis of ALDHbr cells and with standard cell sorters for further purification and characterization.
Brain Tumor Stem Cells, Cancer Cells and Cell Lines, Hematopoietic Stem and Progenitor Cells, Mammary Cells, Mesenchymal Stem and Progenitor Cells, Neural Stem and Progenitor Cells, Other
This product is designed for use in the following research area(s) as part
of the highlighted workflow stage(s). Explore these workflows to learn more about the other products we
offer to support each research area.
The reagents in the kit were frozen when I received it. Will this cause a problem?
No, the reagents in the kit are stable to freezing. Assay performance will not be affected.
Is it acceptable for activation of the ALDEFLUOR™ reagent to exceed 30 minutes?
Yes, as long as room temperature does not exceed 22°C, the reaction can proceed for up to 6 hours with no effect on the assay.
Can I speed up the activation reaction by incubating at 37°C?
This is not recommended. Incubation of the activation reaction at 37°C will not significantly speed up the reaction, and degradation of the activated substrate will occur more quickly at higher temperatures.
Will the activation reaction proceed at refrigerator (2 - 8°C) temperatures?
The ALDEFLUOR™ reagent will remain active for 1 week when stored at 2 - 8°C. For longer storage, divide the remaining reagent into aliquots and store at or below -20°C. Activated ALDEFLUOR™ reagent is stable for 1 year when stored frozen.
How should I store the ALDEFLUOR™ reagent after it is activated?
The ALDEFLUOR™ reagent will remain active for 1 week when stored at 2 - 8°C. For longer storage, divide the remaining reagent into aliquots and store at or below -20°C. Activated ALDEFLUOR™ reagent is stable for 1 year when stored frozen.
Why must the ALDEFLUOR™ assay buffer be added?
This assay has been optimized for detecting stem and progenitor cells by addition of the ALDEFLUOR™ assay buffer. Stem and progenitor cells have high ABC transporter activity and BAAA is a substrate for these efflux pumps. The assay buffer incorporates an efflux pump inhibitor to produce optimal discrimination of the ALDHbr cells and to maximize fluorescent signal stability. We thus recommend that cells be kept on ice and that the ALDEFLUOR™ assay buffer be used throughout all procedures performed after ALDH staining. Not using the assay buffer produces a proportionate loss in the assay signal, depending on the time and temperature at which the stained cells are held.
Is it acceptable for the staining reaction to exceed 30 minutes?
It depends on the cell type. With hematopoietic cells the reaction time can be up to 1 hour at 37°C with no effect on the fluorescence intensity. Incubation periods exceeding 1 hour may lead to an weaker signal and/or higher background. For nonhematopoietic cells optimal incubation times may be different. For example, for the human mammary epithelial SKBR3 cell line, the optimal incubation time was 45 minutes in experiments done at STEMCELL. It is recommended to test different incubation times and determine the optimal incubation time for different cell types.
Will the staining reaction proceed at refrigerator (2 - 8°C) temperatures?
Yes, but full staining will take at least 3 - 4 hours. The staining reaction can continue for up to 24 hours at 2 - 8°C without any effect on the assay.
Can I add any other efflux inhibitors to the ALDEFLUOR™ assay buffer?
Yes. To prevent efflux of the activated ALDEFLUOR™ reagent and the reaction product, the following may be added individually or in combination. These reagents may also improve discrimination of the ALDHbr population, but results will vary by sample type.
• 50 - 100 µM verapamil
• 2.5 mM probenecid
• 100 mM 2-deoxy-D-glucose
• 1 mg/mL sodium azide (0.1%) Note: Sodium azide may be toxic to cells. Do not use if cellular function assays are to be performed after the ALDEFLUOR™ assay.
Note: Ice is the universal efflux inhibitor. Keep all ALDEFLUOR™-reacted samples on ice or at 2 - 8°C as much as possible.
Can I stain the cells at a concentration higher than 1 x 106 cells/mL?
Increasing the concentration of cells up to 5-fold the recommended concentration should have no effect on performance of the assay when using human blood cells. Increasing cell concentrations greater than 5-fold the recommended concentration will decrease assay signal and thereby decrease discrimination of the ALDHbr population. However, different cell types may produce different results. Cell titration experiments may be necessary to determine the optimal cell concentration for different cell types. To stain large number of cells it may be better to increase the sample and reagent volume.
What anticoagulants can be used to collect samples?
Optimal assay performance can be achieved with peripheral blood and leukapheresis samples anticoagulated with acid-citrate dextrose (ACD), ethylenediaminetetraacetic acid (EDTA), or sodium heparin. Bone marrow should be anticoagulated with sodium heparin. Cord blood units may be collected into citrate phosphate dextrose anticoagulant.
Do erythrocytes (red blood cells) interfere with the assay?
The large number of erythrocytes present in peripheral blood, apheresis collections, bone marrow, and umbilical cord blood samples can compete with stem/progenitor cells for the ALDEFLUOR™ substrate. For optimal assay performance, lyse the erythrocytes by treating the samples with ammonium chloride. The ratio of lysis buffer to cell numbers or blood volume must be optimized (10 to 40 parts buffer to sample), and the time (10 - 30 minutes) and temperature (RT or 2 - 8°C) of incubation must be carefully controlled for each lysis buffer and sample type.
What solutions can be used to lyse erythrocytes?
Optimal erythrocyte lysis can be achieved with buffers containing:
• Ammonium chloride (e.g. 0.17 M NH4CI, 10 mM Tris HCI, 0.25 mM EDTA),
• 1X ABC Lysis Buffer (eBioscience, San Diego, CA)
• VitaLyse® (BioE, St Paul, MN).
We do not recommend use of the following or any other solution that contains a fixative, as these will render the cells nonviable:
• CyLyse® (Partec GMBH, Munster, Germany),
• FACS™ Lysing solution (BD Biosciences, San Jose, CA.)
Can fixed cells be used with this assay?
No. The ALDEFLUOR™ reagent is a substrate for the enzyme aldehyde dehydrogenase. ALDEFLUOR™ is a viability marker since the substrate is taken up, catalyzed and retained only by viable cells. It is important to ensure that reagents used for erythrocyte lysis do not contain a fixative.
Does the ALDEFLUOR™ assay work on cryopreserved cells?
ALDEFLUOR™ has been extensively tested on fresh and cryopreserved umbilical cord blood, peripheral blood and leukapheresis samples from patients and mobilized donors. If done correctly, cryopreservation and thawing should not cause loss in cell viability or fluorescence intensity of ALDHbr cells. As only viable cells retain the ALDEFLUOR™ reaction product, a loss in viability will be reflected as a decrease in the percentage of ALDHbr cells and an increase in the percentage of dead/dying cells (detectable by staining for propidium iodide or other viability dyes).
Will ALDEFLUOR™ buffer prevent efflux in cells from non-hematopoietic tissues or from other species?
The proprietary ALDEFLUOR™ assay buffer has been designed to optimize the detection of ALDH-positive (or ALDHbr) cells in human blood. The buffer contains an ATP-binding cassette (ABC) transport inhibitor that prevents active efflux of the ALDEFLUOR™ product from these cells. This transport inhibitor may not prevent efflux from other tissue types or from other species. Consequently, when using samples other than human blood, following the incubation with the activated ALDEFLUOR™ reagent at 37°C, the reacted cells should be kept at 2 - 8°C to prevent efflux, and thus the loss of fluorescence. For a list of additional efflux inhibitors that may be added to the ALDEFLUOR™ buffer see the "CAN I ADD ANY OTHER EFFLUX INHIBITORS TO THE ALDEFLUOR™ ASSAY BUFFER?" question.
Will DEAB inhibit ALDH activity in cells from non-hematopoietic tissues or from other species?
The specific ALDH gene product expressed in non-human, non-blood products may not be inhibited by DEAB. A lack of difference between test and negative control samples may indicate that the inhibitor was not effective, or that there is no ALDH activity in the cells in the sample. Kinetic studies (a progressive increase in ALDEFLUOR™ fluorescence in the negative control tube with time of reaction) may be useful to differentiate these two alternatives. Other ALDH inhibitors can be used as appropriate for the enzyme isoform expressed. For example, Disulfuram inhibits several mammalian ALDH gene products.
Can I use a greater concentration of the ALDEFLUOR™ substrate to improve the discrimination of the ALDHbr population?
When staining non-blood products, it may be necessary to titrate the ALDEFLUOR™ substrate to determine the optimal concentration. We suggest a range of concentrations from 5-fold less to 10-fold more than the standard concentration. During titration we recommend maintaining the concentration of DEAB at 10-fold molar excess of activated ALDEFLUOR™ reagent, and therefore, it is necessary to adjust the amount of DEAB when titrating the substrate.
Can I analyze cells by the ALDEFLUOR™ assay and the side population assay simultaneously?
Yes, the side population assay can be performed in conjunction with the ALDEFLUOR™ assay (Pearce and Bonnet. Exp Hematol 35: 1437-1446, 2007). The Side Population assay should be performed first, followed by the ALDEFLUOR™ assay. We recommend adding 50 µM verapamil to the ALDEFLUOR™ assay buffer when performing both assays.
Why are all the cells in the cytogram fluorescent to some degree?
The ALDEFLUOR™ substrate is a non-polar fluorescent molecule that freely diffuses into all cells. In the DEAB-treated control, fluorescence will reflect the size of the intracellular substrate pool. Fluorescence in the test sample will additionally reflect ALDH activity. Human stem and progenitor cells typically have more ALDH activity than mature cells, and this quantitative difference allows stem cells to be resolved from the other cells.
How do I compensate for multiparameter flow analysis when the staining of ALDHbr cells is so bright?
We would recommend washing your cells with ALDEFLUOR™ assay buffer after the reagent reaction to eliminate background fluorescence from excess substrate. The ALDEFLUOR™ reagent shows an emission spectrum similar to FITC with peak emission at 512 nm. Due to spectral overlap of the ALDEFLUOR™ reagent with fluorochromes that are detected below 650 nm, we recommend using antibodies conjugated to fluorochromes that emit at higher wavelengths for antigens which typically exhibit low levels of expression. For example, when studying the coexpression of CD34 on ALDHbr cells we used the antibody combination, CD45 phycoerythrin (PE), 7- aminoactinomycin D (7-AAD) and CD34 allophycocyanin (APC). Due to the brightness of the ALDEFLUOR™ reagent fluorophore, we strongly recommend the use of compensation controls for every experiment. Adequate compensation will not be achieved with commercially available fluorescent beads.
Diet-Derived Short Chain Fatty Acids Stimulate Intestinal Epithelial Cells To Induce Mucosal Tolerogenic Dendritic Cells.
G. Goverse et al.
Journal of immunology 2017
Abstract
The gastrointestinal tract is continuously exposed to many environmental factors that influence intestinal epithelial cells and the underlying mucosal immune system. In this article, we demonstrate that dietary fiber and short chain fatty acids (SCFAs) induced the expression of the vitamin A-converting enzyme RALDH1 in intestinal epithelial cells in vivo and in vitro, respectively. Furthermore, our data showed that the expression levels of RALDH1 in small intestinal epithelial cells correlated with the activity of vitamin A-converting enzymes in mesenteric lymph node dendritic cells, along with increased numbers of intestinal regulatory T cells and a higher production of luminal IgA. Moreover, we show that the consumption of dietary fiber can alter the composition of SCFA-producing microbiota and SCFA production in the small intestines. In conclusion, our data illustrate that dietary adjustments affect small intestinal epithelial cells and can be used to modulate the mucosal immune system.
MSCs derived from iPSCs with a modified protocol are tumor-tropic but have much less potential to promote tumors than bone marrow MSCs.
Zhao Q et al.
Proceedings of the National Academy of Sciences of the United States of America 2015 JAN
Abstract
Mesenchymal stem or stromal cells (MSCs) have many potential therapeutic applications including therapies for cancers and tissue damages caused by cancers or radical cancer treatments. However, tissue-derived MSCs such as bone marrow MSCs (BM-MSCs) may promote cancer progression and have considerable donor variations and limited expandability. These issues hinder the potential applications of MSCs, especially those in cancer patients. To circumvent these issues, we derived MSCs from transgene-free human induced pluripotent stem cells (iPSCs) efficiently with a modified protocol that eliminated the need of flow cytometric sorting. Our iPSC-derived MSCs were readily expandable, but still underwent senescence after prolonged culture and did not form teratomas. These iPSC-derived MSCs homed to cancers with efficiencies similar to BM-MSCs but were much less prone than BM-MSCs to promote the epithelial-mesenchymal transition, invasion, stemness, and growth of cancer cells. The observations were probably explained by the much lower expression of receptors for interleukin-1 and TGFβ, downstream protumor factors, and hyaluronan and its cofactor TSG6, which all contribute to the protumor effects of BM-MSCs. The data suggest that iPSC-derived MSCs prepared with the modified protocol are a safer and better alternative to BM-MSCs for therapeutic applications in cancer patients. The protocol is scalable and can be used to prepare the large number of cells required for off-the-shelf" therapies and bioengineering applications."
Neutrophils support lung colonization of metastasis-initiating breast cancer cells
Wculek SK and Malanchi I
Nature 2015 DEC
Abstract
Despite progress in the development of drugs that efficiently target cancer cells, treatments for metastatic tumours are often ineffective. The now well-established dependency of cancer cells on their microenvironment suggests that targeting the non-cancer-cell component of the tumour might form a basis for the development of novel therapeutic approaches. However, the as-yet poorly characterized contribution of host responses during tumour growth and metastatic progression represents a limitation to exploiting this approach. Here we identify neutrophils as the main component and driver of metastatic establishment within the (pre-)metastatic lung microenvironment in mouse breast cancer models. Neutrophils have a fundamental role in inflammatory responses and their contribution to tumorigenesis is still controversial. Using various strategies to block neutrophil recruitment to the pre-metastatic site, we demonstrate that neutrophils specifically support metastatic initiation. Importantly, we find that neutrophil-derived leukotrienes aid the colonization of distant tissues by selectively expanding the sub-pool of cancer cells that retain high tumorigenic potential. Genetic or pharmacological inhibition of the leukotriene-generating enzyme arachidonate 5-lipoxygenase (Alox5) abrogates neutrophil pro-metastatic activity and consequently reduces metastasis. Our results reveal the efficacy of using targeted therapy against a specific tumour microenvironment component and indicate that neutrophil Alox5 inhibition may limit metastatic progression.
New look, same high quality and support! You may notice that your instrument or reagent packaging looks slightly different from images displayed on the website, or from previous orders. We are updating our look but rest assured, the products themselves and how you should use them have not changed. Learn more
Legal Statement:
ALDEFLUOR is registered trademark of ALDAGEN Inc.
Quality Statement:
PRODUCTS ARE FOR RESEARCH USE ONLY AND NOT INTENDED FOR HUMAN OR ANIMAL DIAGNOSTIC OR THERAPEUTIC USES UNLESS OTHERWISE STATED. FOR ADDITIONAL INFORMATION ON QUALITY AT STEMCELL, REFER TO WWW.STEMCELL.COM/COMPLIANCE.