Make more informed purchasing decisions with our new product availability and delivery estimate feature, now available on all product pages, in your cart, and during checkout.
Sign In
New to STEMCELL?
Register for an account to quickly and easily purchase products online and for one-click access to all educational content.
Products sold in North America are collected in the U.S. and Institutional Review Board (IRB)-approved. Products sold in the U.K. and EU are collected in the U.K. at a facility licensed by the HTA (Human Tissue Authority) and using consent forms and protocols approved by a Research Ethics Committee (REC).
Request Pricing
Thank you for your interest in this product.
Please provide us with your contact information and your local representative
will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration
By submitting this form, you are providing your consent to STEMCELL Technologies Canada Inc. and its subsidiaries and affiliates (“STEMCELL”) to collect and use your information, and send you newsletters and emails in accordance with our privacy policy. Please contact us with any questions that you may have. You can unsubscribe or change your email preferences at any time.
Obtain large numbers of single-donor mononuclear cells (PBMCs) from normal peripheral blood and reduce the time and reagents required for your downstream cell isolation. We help you get the cells you need with personalized service, custom products, flexible delivery times, and multiple size options.
Leukopaks are highly concentrated, low-volume apheresis collections from normal donors, with with Institutional Review Board (IRB)- or Research Ethics Committee (REC)-approved consent forms and protocols. Each full-sized leukopak is produced from ~2 - 3x blood volumes using the Spectra Optia® Apheresis System, with acid-citrate-dextrose solution A (ACDA) as the anticoagulant. High-resolution HLA typing (Class I and Class II alleles and CMV status) are available upon request. Donor specifications (e.g. BMI category, smoking status, ethnicity, etc.) can be requested in the comment box above, after selecting from the product options. Products sold in North America are collected in the U.S. and Institutional Review Board (IRB)-approved. Products sold in the U.K. and EU are collected in the U.K. at a facility licensed by the HTA (Human Tissue Authority) and using consent forms and protocols approved by a Research Ethics Committee (REC). U.S. donors are screened for HIV-1, HIV-2, hepatitis B, and hepatitis C, and U.K. donors are additionally screened for HTLV I/II and syphilis.
Certain products are only available in select territories. Please contact your sales representative or Product & Scientific Support at techsupport@stemcell.com for further information.
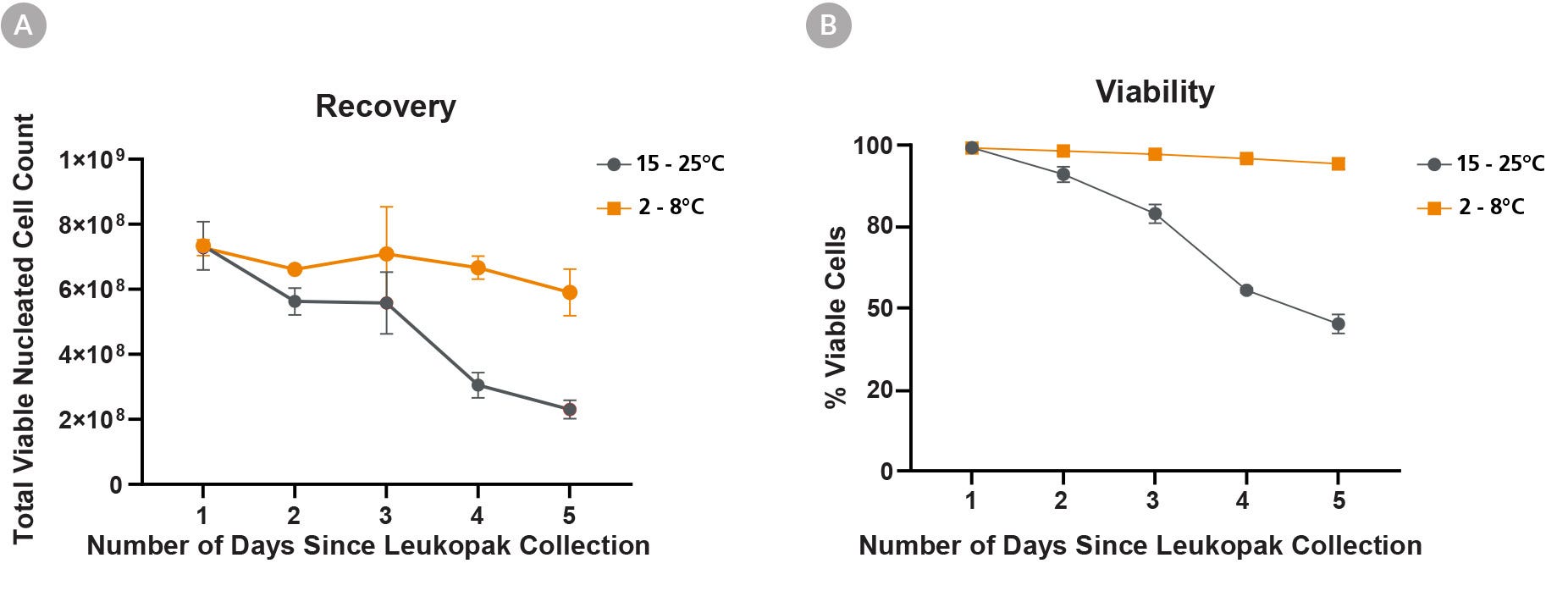
Figure 1. Recovery of Viable Cells Is Higher When Leukopaks Are Stored at Fridge Temperature Compared to Room Temperature
Stability of fresh leukopaks (LP) was evaluated after storage for up to 5 days after collection, at a refrigerated temperature (“FT”; 2 - 8°C) or at room temperature (RT; 15 - 25°C). Full leukopaks were collected from three (3) unique donors, then divided into ten (10) equally sized fractions and stored at FT or RT (n = 5 at each temp, per donor). Over the following 5 days, one (1) 1/10th leukopak fraction was processed from each condition daily, to identify changes in cellular composition and functionality that may have occurred during storage.
Viable cell yield (A) and percent cell viability (B) were determined daily, following red blood cell (RBC) lysis of leukopak fractions. By day 5 after collection, leukopak fractions stored at RT showed a 68% loss of viable cells and an accompanying 54% decrease in cell viability. Conversely, those stored at FT showed a much less pronounced 19% decrease of viable cells and just 5% decrease in viability over the 5 days. All data points represent average ± standard deviation values from leukopak fractions of 3 unique donors.
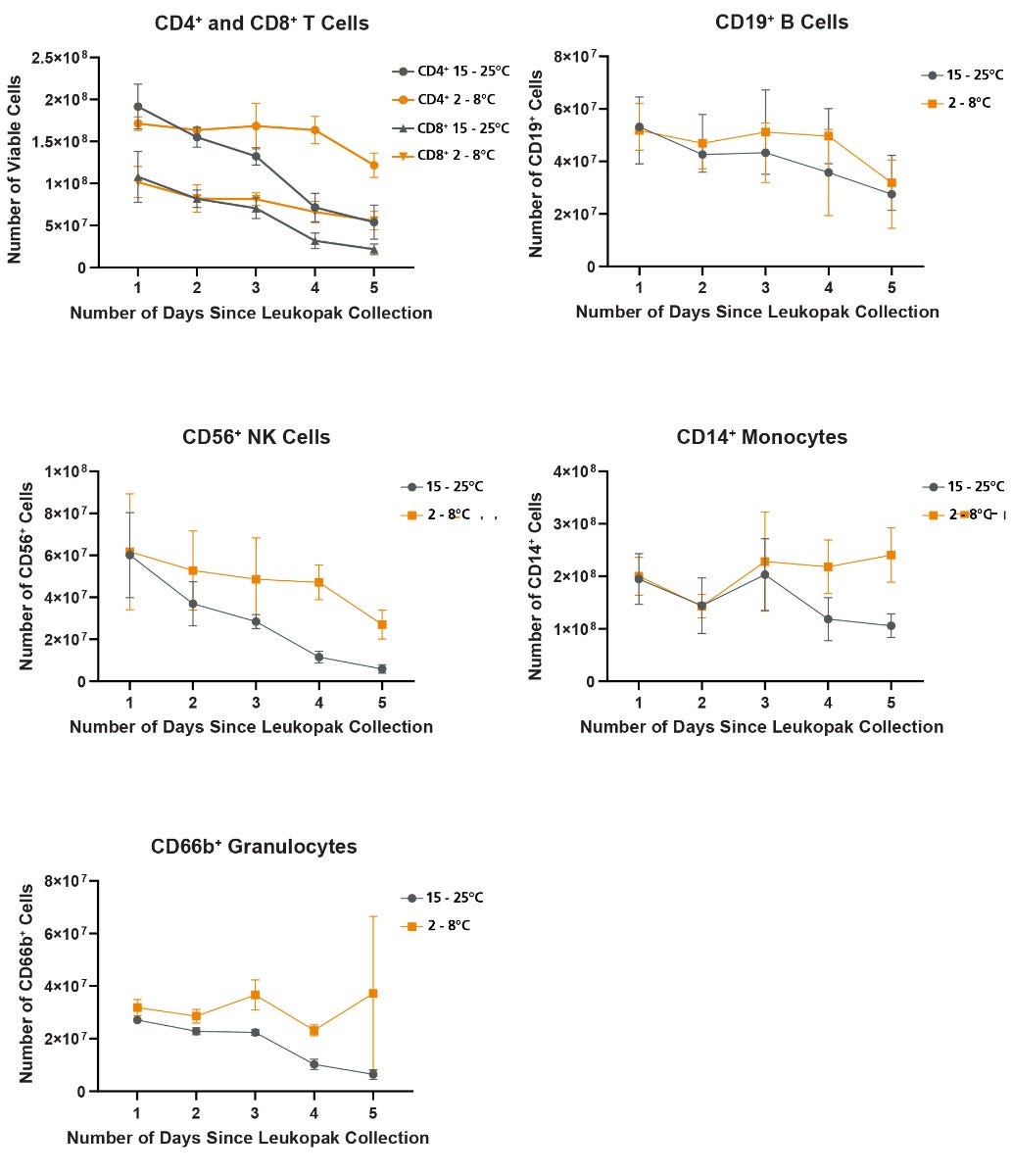
Figure 2. Refrigeration of Leukopaks Ameliorates Time-Dependent Loss of Mature Blood Cells
Recovery of mature blood cells from leukopaks stored at RT or FT was calculated from the viable total cell recovery combined with lineage frequency assessment by flow cytometry. For all mature cell types examined, recovery decreased at a faster rate when leukopaks were stored at RT compared to FT. A pronounced loss of T and NK cells was particularly evident over the 5 days in leukopaks stored at RT. Less pronounced losses of B cells, monocytes and granulocytes were also observed, and these losses were also ameliorated by FT storage. All data points represent average ± standard deviation values from leukopak fractions of n = 3 unique donors.
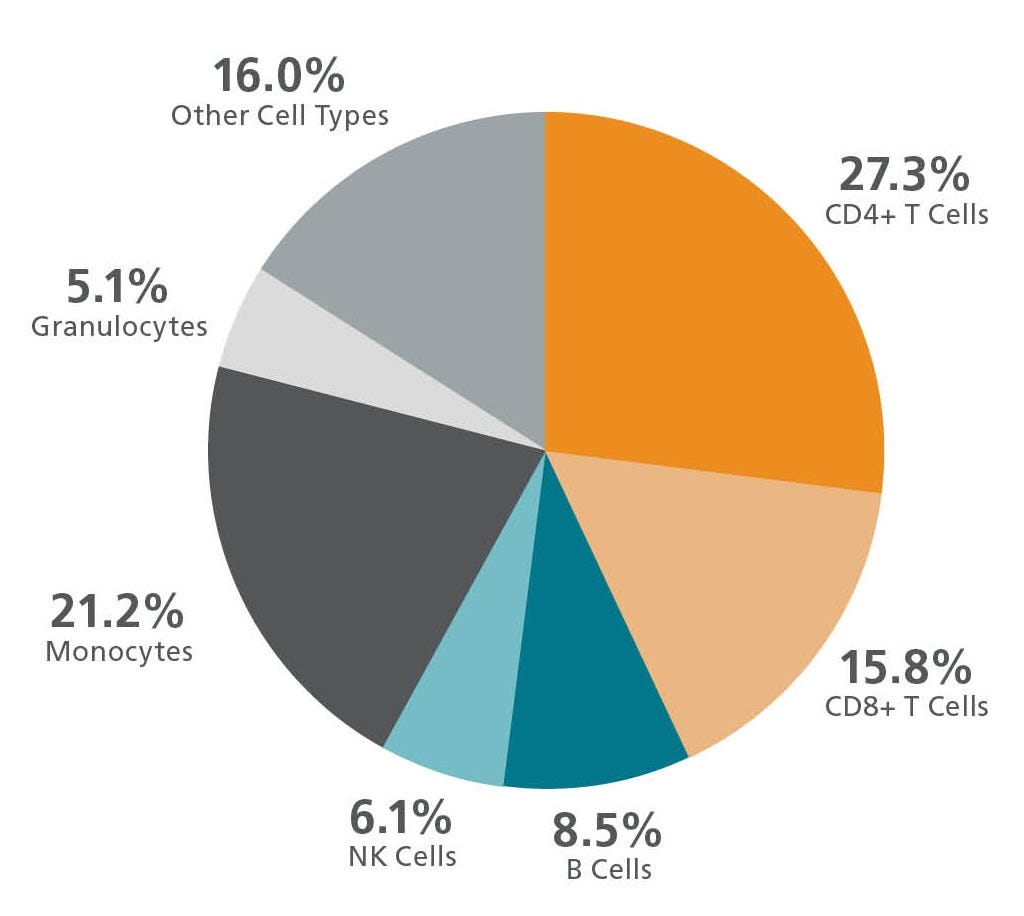
Figure 3. Mean Percentages of Cell Subpopulations in a Leukopak
Representative chart showing the average frequencies of major immune subsets in leukopak products, as measured by flow cytometry. Values shown are mean percentages of total viable leukocytes present in a leukopak (n ≥ 30).
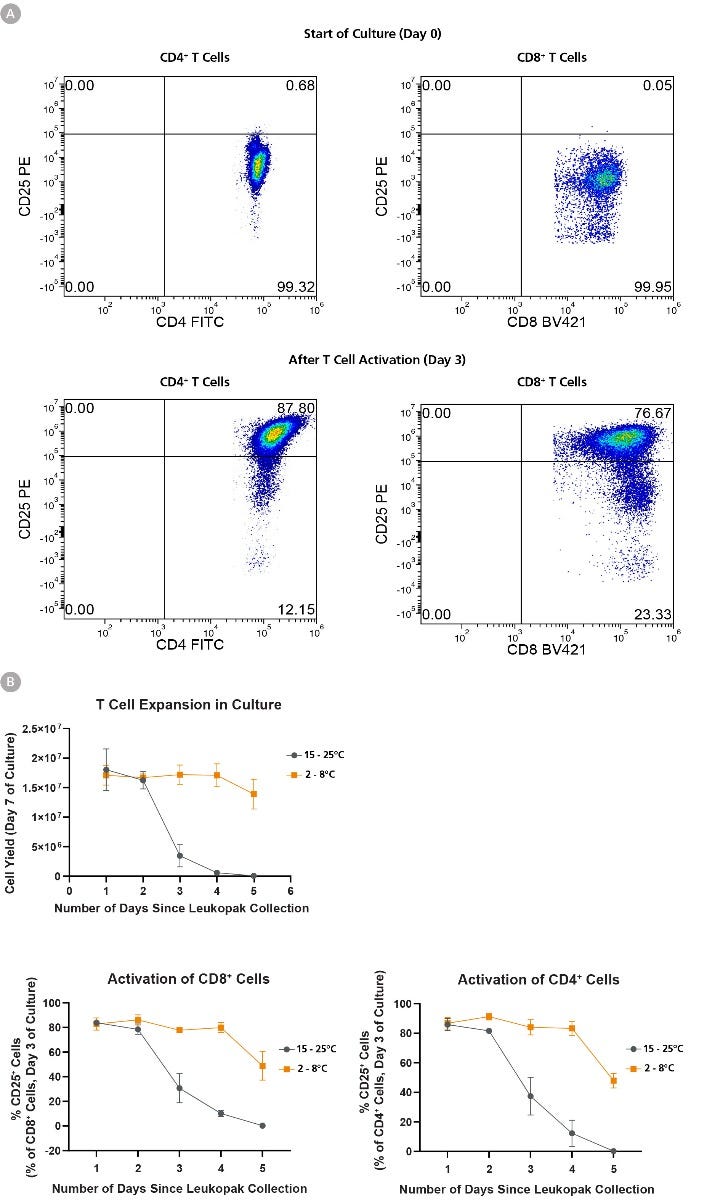
Figure 4. T Cell Expansion and Activation Ability Is Better Preserved When Leukopaks are Stored at Fridge Temperature
Using EasySep™ Human T Cell Isolation Kit, T cells were isolated from 1 leukopak fraction of each storage condition daily for 5 days, and 1 x 10^6 isolated cells were cultured in ImmunoCult™-XF T Cell Expansion Medium supplemented with ImmunoCult™ Human CD3/CD28/CD2 T Cell Activator and 10 ng/mL IL-2 for 10 days with assessment of CD25 activation marker expression on day 3 of culture. (A) Representative flow cytometry data from leukopaks stored 1 day at FT, showing that both CD4+ and CD8+ cells are CD25-negative at the start of culture (day 0), and upregulate CD25 expression by day 3 of culture. (B) Cellular expansion and corresponding cell yield over 7 days of culture decreases in correlation with storage duration of leukopak fractions. Leukopaks that were stored for 1 - 2 days at either RT or FT had high expansion potential yielding 1.5 - 2 x 10^7 cells, and this 15 - 20-fold expansion potential is maintained in T cells from leukopaks stored at FT for up to 5 days. In contrast, little or no expansion is observed in T cell cultures from leukopaks stored at RT for 3 or more days, indicating a loss of proliferative capacity. Moreover, T cells show a gradual reduction in their ability to become activated by ImmunoCult™ T Cell activator, as shown by a reduction in day 3 CD25 expression in gated CD4+ (bottom left) or CD8+ (bottom right) cells over time, and the effect is most pronounced with storage at RT for 3 or more days. All data points represent average ± standard deviation values from leukopak fractions of n = 3 unique donors.
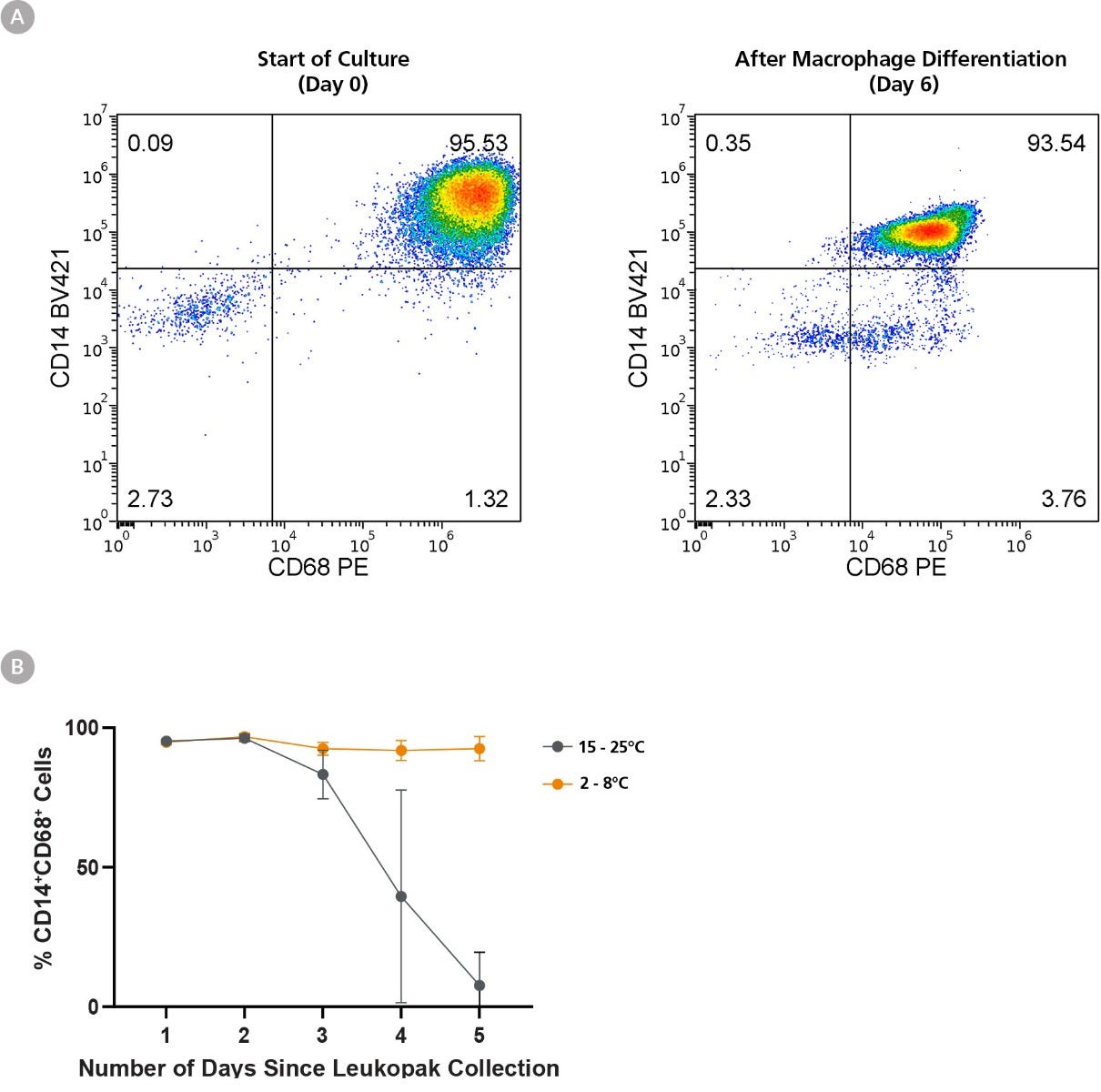
Figure 5. Refrigeration of Leukopaks Preserves Monocyte-to-Macrophage Differentiation Efficiency for Up to 5 Days
Monocytes were isolated (using EasySep™ Human Monocyte Isolation Kit) from 1 leukopak fraction of each storage condition daily for 5 days, and 1 x 10^6 isolated cells were cultured in ImmunoCult™-SF Macrophage Medium supplemented with 50 ng/mL Human Recombinant M-CSF for a further 6 days. (A) Representative flow cytometry plot from leukopaks stored 1 day at FT, showing maintenance of CD14 and upregulation of CD68 expression over the 5 day differentiation to M0 macrophages. (B) While monocytes isolated from FT-stored leukopaks efficiently differentiated into M0 macrophages, those stored at RT for over 3 days failed to differentiate, as shown by low percentages of CD14+CD68+ cells. Moreover, monocytes harvested from Day 5 leukopaks stored at RT failed to thrive in the 6-day culture, resulting in very few viable cells recovered (not shown). All data points represent average ± standard deviation values from leukopak fractions of n = 3 unique donors.
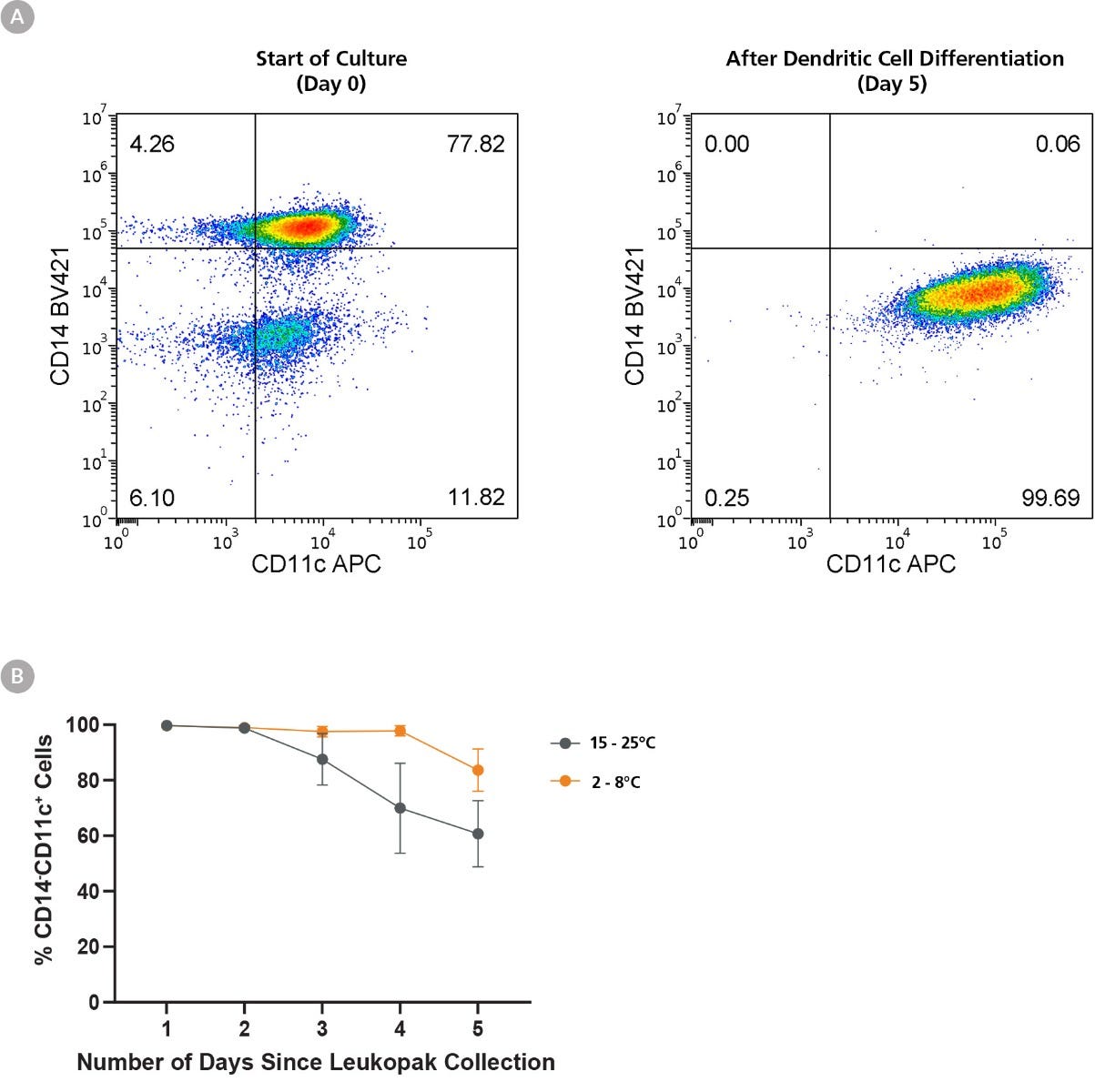
Figure 6. Refrigeration of Leukopaks Preserves Monocyte-to-Dendritic Cell Differentiation Ability for Up to 5 Days
Monocytes were isolated (using EasySep™ Human Monocyte Isolation Kit) from 1 leukopak fraction of each storage condition daily for 5 days, and 1 x 10^6 isolated cells were cultured in ImmunoCult™-ACF Dendritic Cell (DC) Medium supplemented with ImmunoCult™-ACF Dendritic Cell Differentiation Supplement. (A) Representative flow cytometry plot from leukopaks stored 1 day at FT, showing efficient downregulation of CD14 and upregulation of CD11c over 6 days of culture. (B) While monocytes isolated from FT-stored leukopaks efficiently differentiated into DCs, those stored at RT for over 3 days show reduced DC differentiation ability and CD14-CD11c+ cell output. Moreover, monocytes harvested from day 5 RT-stored leukopaks failed to thrive in the 5-day culture, resulting in very few viable cells recovered (not shown). All data points represent average ± standard deviation values from leukopak fractions of n = 3 unique donors.
This product is designed for use in the following research area(s) as part
of the highlighted workflow stage(s). Explore these workflows to learn more about the other products we
offer to support each research area.
Gut-Liver Physiomimetics Reveal Paradoxical Modulation of IBD-Related Inflammation by Short-Chain Fatty Acids.
M. Trapecar et al.
Cell systems 2020 mar
Abstract
Although the association between the microbiome and IBD and liver diseases is known, the cause and effect remain elusive. By connecting human microphysiological systems of the gut, liver, and circulating Treg and Th17 cells, we created a multi-organ model of ulcerative colitis (UC) ex vivo. The approach shows microbiome-derived short-chain fatty acids (SCFAs) to either improve or worsen UC severity, depending on the involvement of effector CD4 T cells. Using multiomics, we found SCFAs increased production of ketone bodies, glycolysis, and lipogenesis, while markedly reducing innate immune activation of the UC gut. However, during acute T cell-mediated inflammation, SCFAs exacerbated CD4+ T cell-effector function, partially through metabolic reprograming, leading to gut barrier disruption and hepatic injury. These paradoxical findings underscore the emerging utility of human physiomimetic technology in combination with systems immunology to study causality and the fundamental entanglement of immunity, metabolism, and tissue homeostasis.
Impact of mRNA chemistry and manufacturing process on innate immune activation.
J. Nelson et al.
Science advances 2020 jun
Abstract
Messenger RNA (mRNA) represents an attractive therapeutic modality for potentially a wide range of clinical indications but requires uridine chemistry modification and/or tuning of the production process to prevent activation of cellular innate immune sensors and a concomitant reduction in protein expression. To decipher the relative contributions of these factors on immune activation, here, we compared, in multiple cell and in vivo models, mRNA that encodes human erythropoietin incorporating either canonical uridine or N1-methyl-pseudouridine (1m$\Psi$), synthesized by either a standard process shown to have double-stranded RNA (dsRNA) impurities or a modified process that yields a highly purified mRNA preparation. Our data demonstrate that the lowest stimulation of immune endpoints was with 1m$\Psi$ made by the modified process, while mRNA containing canonical uridine was immunostimulatory regardless of process. These findings confirm that uridine modification and the reduction of dsRNA impurities are both necessary and sufficient at controlling the immune-activating profile of therapeutic mRNA.
Integrated gut/liver microphysiological systems elucidates inflammatory inter-tissue crosstalk.
Chen WLK et al.
Biotechnology and bioengineering 2017
Abstract
A capability for analyzing complex cellular communication among tissues is important in drug discovery and development, and in vitro technologies for doing so are required for human applications. A prominent instance is communication between the gut and the liver, whereby perturbations of one tissue can influence behavior of the other. Here, we present a study on human gut-liver tissue interactions under normal and inflammatory contexts, via an integrative multi-organ platform comprising human liver (hepatocytes and Kupffer cells), and intestinal (enterocytes, goblet cells, and dendritic cells) models. Our results demonstrated long-term (>2 weeks) maintenance of intestinal (e.g., barrier integrity) and hepatic (e.g., albumin) functions in baseline interaction. Gene expression data comparing liver in interaction with gut, versus isolation, revealed modulation of bile acid metabolism. Intestinal FGF19 secretion and associated inhibition of hepatic CYP7A1 expression provided evidence of physiologically relevant gut-liver crosstalk. Moreover, significant non-linear modulation of cytokine responses was observed under inflammatory gut-liver interaction; for example, production of CXCR3 ligands (CXCL9,10,11) was synergistically enhanced. RNA-seq analysis revealed significant upregulation of IFNα/β/γ signaling during inflammatory gut-liver crosstalk, with these pathways implicated in the synergistic CXCR3 chemokine production. Exacerbated inflammatory response in gut-liver interaction also negatively affected tissue-specific functions (e.g., liver metabolism). These findings illustrate how an integrated multi-tissue platform can generate insights useful for understanding complex pathophysiological processes such as inflammatory organ crosstalk. Biotechnol. Bioeng. 2017;114: 2648-2659. textcopyright 2017 Wiley Periodicals, Inc.
Products sold in North America are collected in the U.S. and Institutional Review Board (IRB)-approved. Products sold in the U.K. and EU are collected in the U.K. at a facility licensed by the HTA (Human Tissue Authority) and using consent forms and protocols approved by a Research Ethics Committee (REC).
Quality Statement:
PRODUCTS ARE FOR RESEARCH USE ONLY AND NOT INTENDED FOR HUMAN OR ANIMAL DIAGNOSTIC OR THERAPEUTIC USES UNLESS OTHERWISE STATED.