Make more informed purchasing decisions with our new product availability and delivery estimate feature, now available on all product pages, in your cart, and during checkout.
Sign In
New to STEMCELL?
Register for an account to quickly and easily purchase products online and for one-click access to all educational content.
Thank you for your interest in this product.
Please provide us with your contact information and your local representative
will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration
By submitting this form, you are providing your consent to STEMCELL Technologies Canada Inc. and its subsidiaries and affiliates (“STEMCELL”) to collect and use your information, and send you newsletters and emails in accordance with our privacy policy. Please contact us with any questions that you may have. You can unsubscribe or change your email preferences at any time.
StemSpan™ Serum-Free Expansion Medium (SFEM) has been developed and tested for the in vitro culture and expansion of human hematopoietic cells, when the appropriate growth factors and supplements are added. This allows users the flexibility to prepare medium that meets their requirements. When combined with the appropriate cytokines, SFEM has been used for the culture and expansion of hematopoietic cells isolated from other species, including mouse, non-human primate, and dog. SFEM has also been used for culture of various other hematopoietic and non-hematopoietic cell types. Using appropriate StemSpan™ Expansion Supplements, SFEM may be used to expand CD34+ cells isolated from human cord blood, mobilized peripheral blood, or bone marrow samples, or to expand and differentiate lineage-committed progenitors to generate populations of erythroid, myeloid, or megakaryocyte progenitor cells.
StemSpan™ SFEM II (Catalog #09605) is an improved version of StemSpan™ SFEM that is further enriched to promote and support higher rates of CD34+ expansion and/or cell differentiation.
Contains
• Iscove’s MDM
• Bovine serum albumin
• Recombinant human insulin
• Human transferrin (iron-saturated)
• 2-Mercaptoethanol
• Supplements
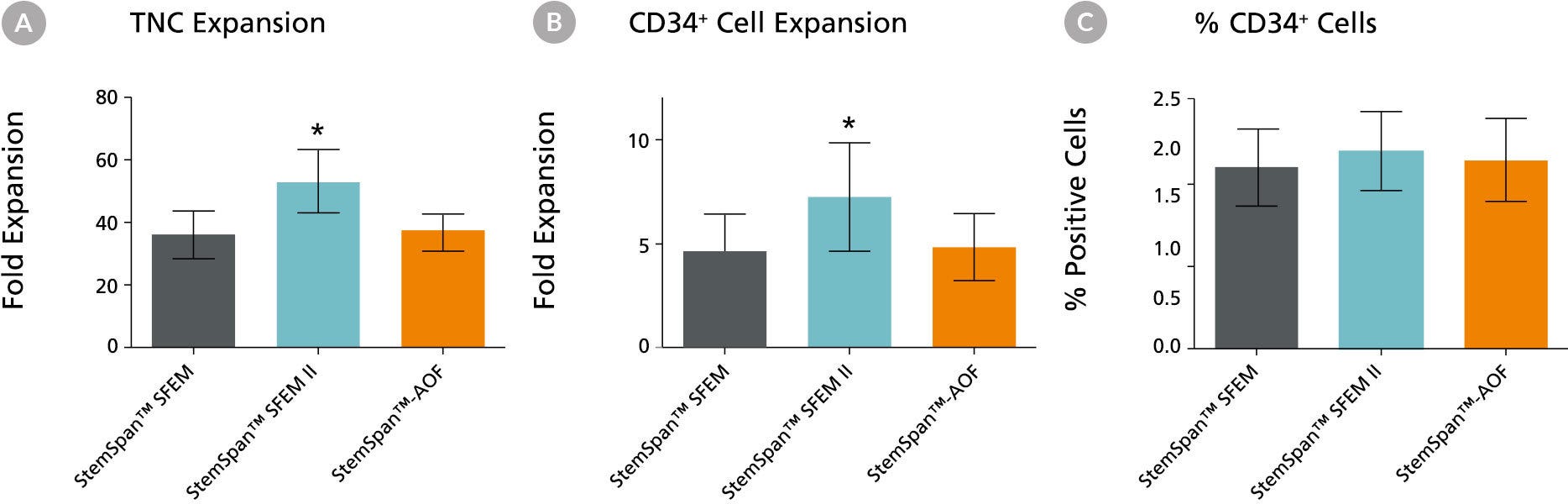
Figure 1. Expansion of CD34+ Human Cord Blood Cells Cultured in StemSpan™ Media Containing CC100 Cytokine Cocktail
Purified CD34+ human cord blood (CB) cells were suspended at a concentration of 10,000 per mL in StemSpan™ SFEM (dark gray bars), SFEM II (blue bars) and AOF (orange bars) media containing CC100 Cytokine Cocktail (Catalog #02690). Cultures were maintained for 7 days, after which the cells were counted and examined for CD34 and CD45 expression by flow cytometry. Shown are the fold expansion of total nucleated cells (TNC) (A) and CD34+ cells (B) per input CD34+ cell, and the percent CD34 + cells (C). Results represent the average results of 32 different CB samples. Vertical lines indicate 95% confidence limits, the range within which 95% of results fall. The numbers of cells produced in StemSpan™ SFEM II were significantly higher than in StemSpan™ SFEM and StemSpan™-AOF (*p<0.001, paired t-test, n=32).
Note: Data for StemSpan™-AOF shown were generated with the original phenol red-containing version StemSpan™-ACF (Catalog #09855). However internal testing showed that the performance of the new phenol red-free, cGMP-manufactured version, StemSpan™-AOF (Catalog #100-0130) was comparable.
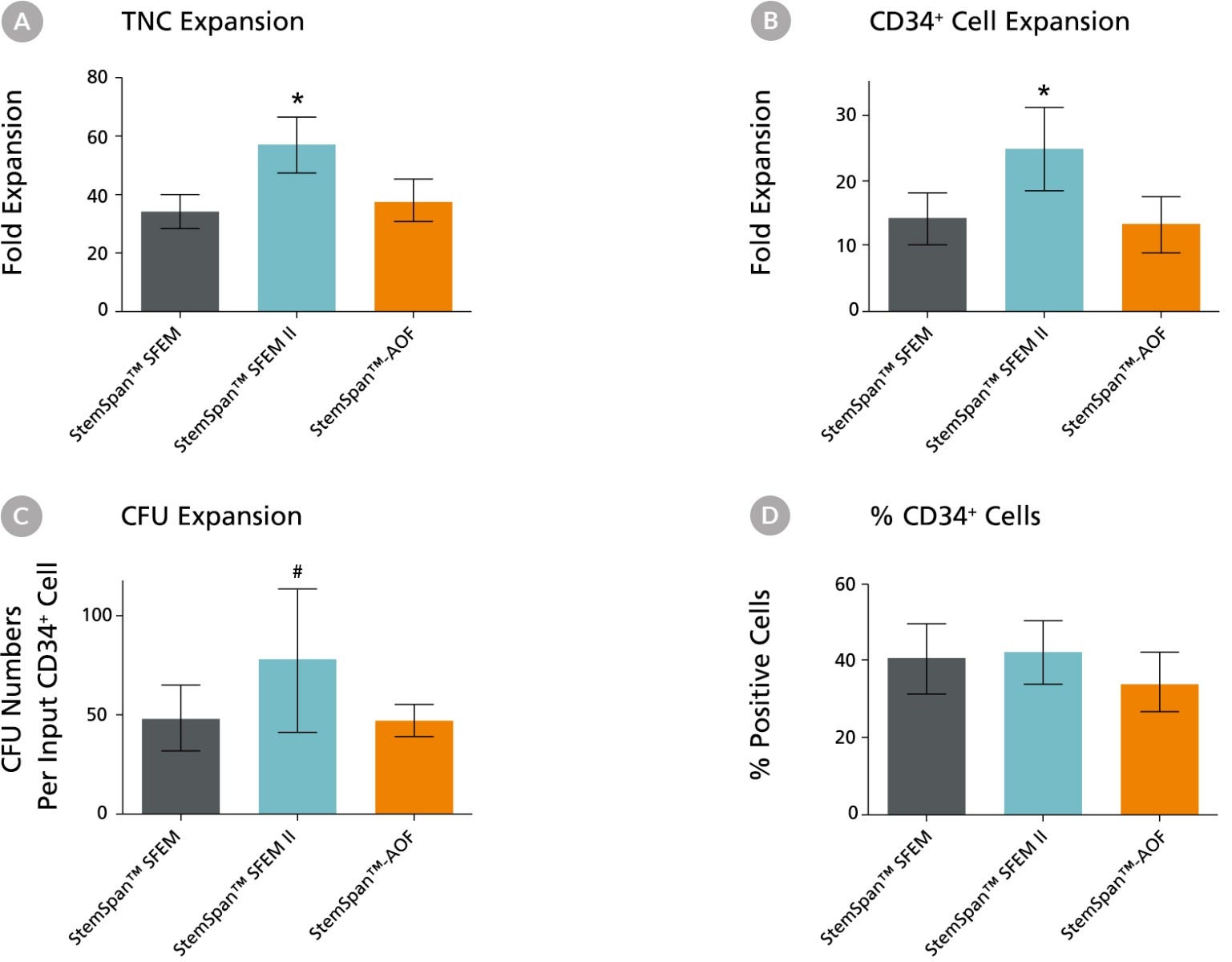
Figure 2. Expansion of CD34+ Human Cord Blood Cells Cultured in StemSpan™ Media Containing CD34+ Expansion Supplement
Purified CD34+ human cord blood (CB) cells were suspended at a concentration of 10,000 per mL in StemSpan™ SFEM (dark gray bars), SFEM II (blue bars) and AOF (orange bars) media containing CD34+ Expansion Supplement (Catalog #02691). Cultures were maintained for 7 days, after which the cells were counted and examined for CD34 and CD45 expression by flow cytometry. The number of colony-forming units (CFU) in the expanded population was determined by replating cells in MethoCult™ H4435 and counting the number of colonies produced 14 days later. Shown are the fold expansion of total nucleated cells (TNC) (A), CD34+ cells (B) and CFU numbers (C) per input CD34+ cell, and the percent CD34+ cells (D) in these cultures (n=6). Vertical lines indicate 95% confidence limits, the range within which 95% of results fall. The numbers of cells produced in StemSpan™ SFEM II was significantly higher than in SFEM and AOF (*p<0.001, #p<0.05, paired t-test, n=6).
Note: Data for StemSpan™-AOF shown were generated with the original phenol red-containing version StemSpan™-ACF (Catalog #09855). However internal testing showed that the performance of the new phenol red-free, cGMP-manufactured version, StemSpan™-AOF (Catalog #100-0130) was comparable.
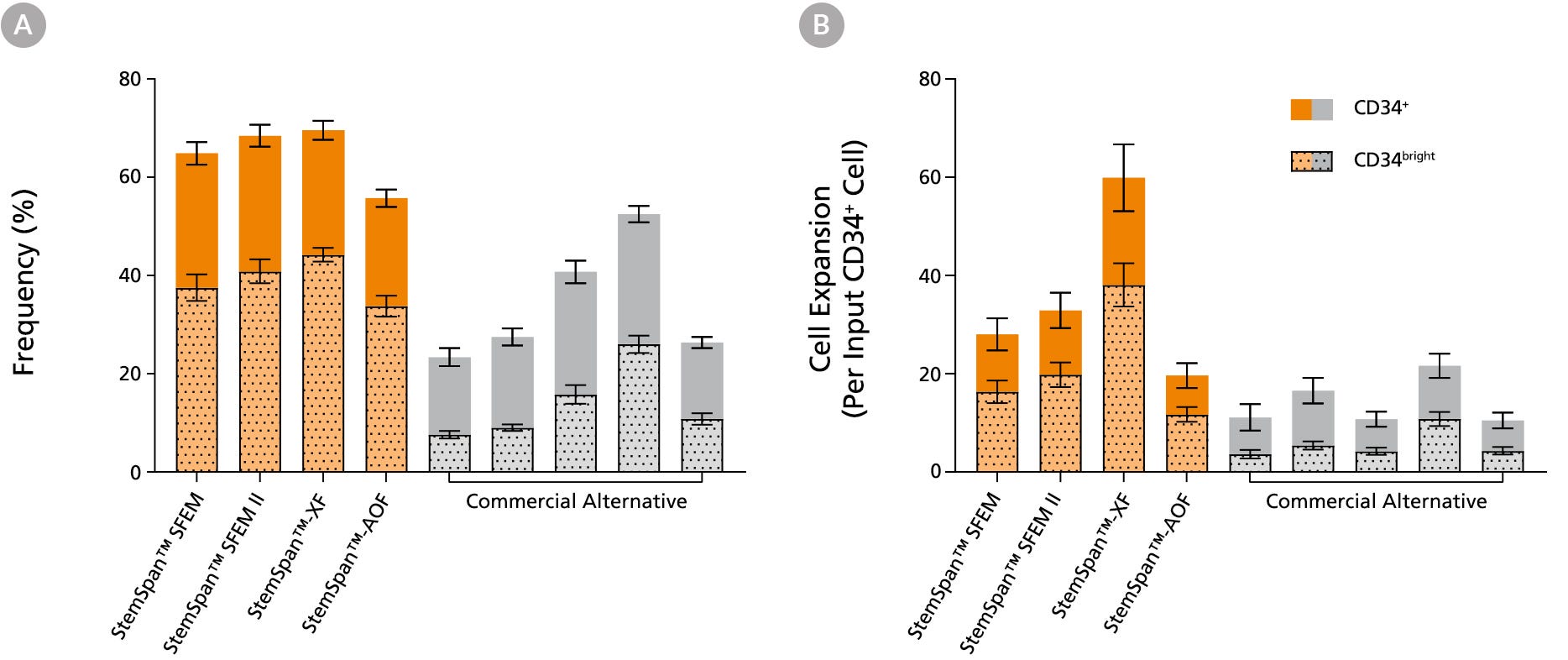
Figure 3. StemSpan™ Media Support Greater Expansion of Human CD34+ and CD34bright Cells than Other Commercial Media
Purified CB-derived CD34+ cells were cultured for 7 days in select StemSpan™ media (StemSpan™ SFEM, StemSpan™ SFEM II, StemSpan™-XF, or StemSpan™-AOF, orange bars), and in five xeno-free media formulations from other suppliers (Xeno-Free Commercial Alternative, grey bars) including (in random order) CTS™ StemPro™ HSC (Thermo), SCGM (Cellgenix), X-VIVO™ 15 (Lonza), Stemline™ II (Sigma), and StemPro™-34 (Thermo). All media were supplemented with StemSpan™ CD34+ Expansion Supplement and UM171*. The (A) frequency and (B) cell expansion of viable CD34+ and CD34bright cells in culture were based on viable cell counts and flow cytometry results as shown in Figure 1. StemSpan™ showed significantly higher expansion of CD34+ and CD34bright cells (P < 0.05 when comparing StemSpan™ SFEM II to five media from other suppliers, calculated using a one-way ANOVA followed by Dunnett’s post hoc test) and StemSpan™-AOF, the only animal origin-free formulation, showed equivalent performance to all xeno-free commericals alternatives tested. Data shown are mean ± SEM (n = 8).
Note: Data for StemSpan™-AOF shown were generated with the original phenol red-containing version StemSpan™-ACF (Catalog #09855). However internal testing showed that the performance of the new phenol red-free, cGMP-manufactured version, StemSpan™-AOF (Catalog #100-0130) was comparable.
*Similar results are expected when using UM729 (Catalog #72332) prepared to a final concentration of 1μM. For more information including data comparing UM171 and UM729, see Fares et al., 2014.
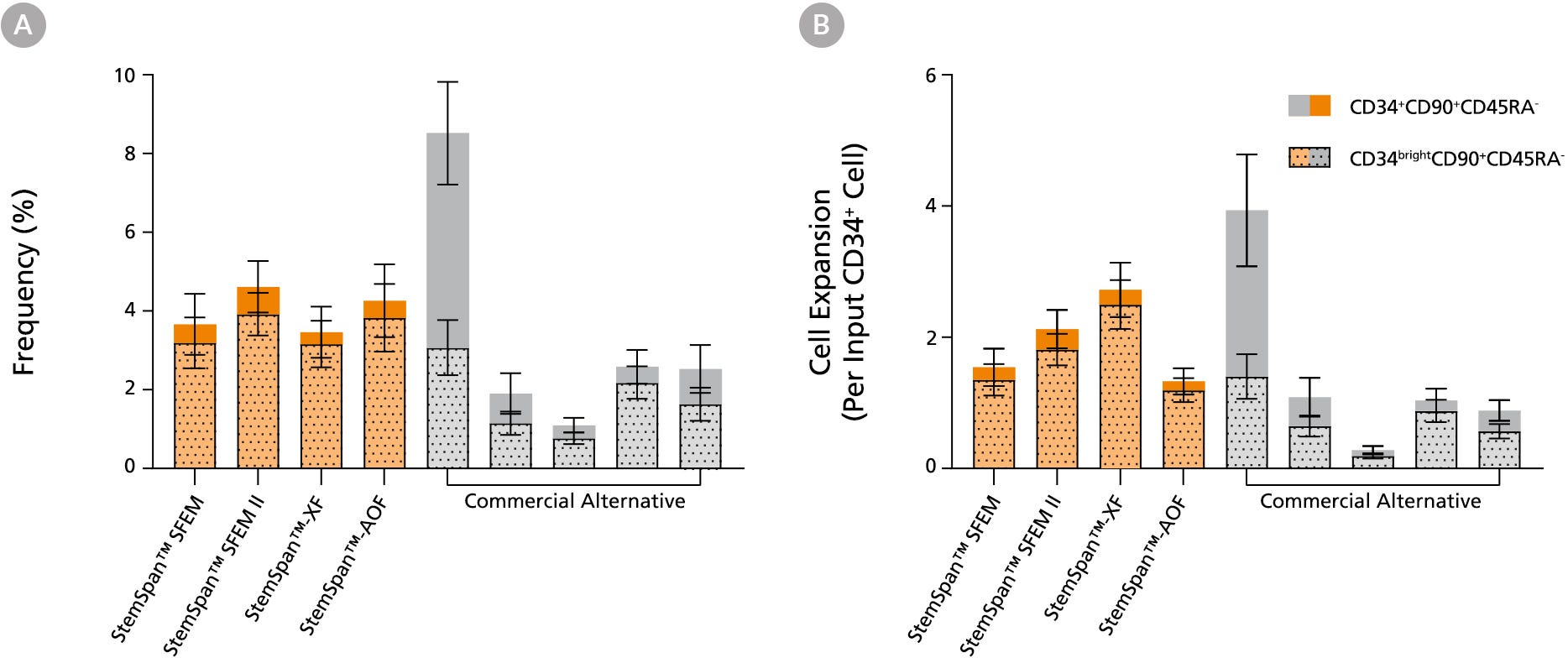
Figure 4. StemSpan™ Media Support Equal or Greater Expansion of Primitive Human CD34brightCD90+CD45RA- Cells Than Other Commercial Media
Purified CB-derived CD34+ cells were cultured for 7 days in select StemSpan™ media (StemSpan™ SFEM, StemSpan™ SFEM II, StemSpan™-XF, or StemSpan™-AOF, orange bars), and in five xeno-free media formulations from other suppliers (Commercial Alternative, grey bars) including (in random order) CTS StemPro HSC (Thermo), SCGM (Cellgenix), X-VIVO 15 (Lonza), Stemline II (Sigma), and StemPro 34 (Thermo). All media were supplemented with StemSpan™ CD34+ Expansion Supplement and UM171*. The (A) frequency and (B) cell expansion of CD34+CD90+CD45RA- (solid) and CD34brightCD90+CD45RA-(dotted overlay) cells in culture were based on viable cell counts and flow cytometry results as shown in Figure 1. StemSpan™ media showed similar or significantly higher expansion of CD34brightCD90+CD45RA- cells (P < 0.05 compared to five media from other suppliers, calculated using one-way ANOVA followed by Dunnett’s post hoc test) and StemSpan™-AOF, the only animal origin-free formulation tested, showed equivalent performance to all xeno-free commercial alternatives tested. Data shown are mean ± SEM (n = 8).
Note: Data for StemSpan™-AOF shown were generated with the original phenol red-containing version StemSpan™-ACF (Catalog #09855). However internal testing showed that the performance of the new phenol red-free, cGMP-manufactured version, StemSpan™-AOF (Catalog #100-0130) was comparable.
*Similar results are expected when using UM729 (Catalog #72332) prepared to a final concentration of 1μM. For more information including data comparing UM171 and UM729, see Fares et al. 2014.
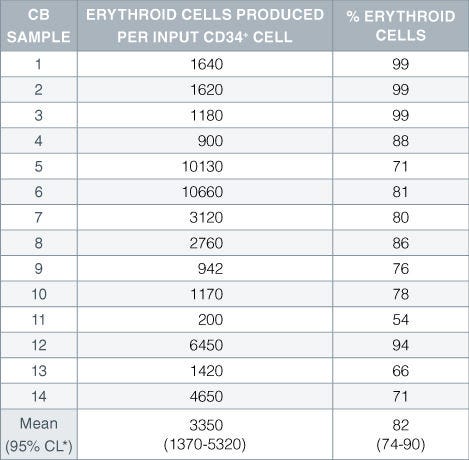
Table 1. Production of Erythroid Cells From CD34+ Human Cord Blood Cells Cultured in StemSpan™ SFEM Serum-Free Expansion Medium Containing Erythroid Expansion Supplement
Numbers and percent of erythroid cells produced after 14 days of culture of enriched CD34 + cells from 14 different cord blood (CB) samples. Erythroid cells were characterized by flow cytometry on the basis of transferrin receptor (CD71) and glycophorin A (CD235) expression.*95% confidence limits, the range within which 95% of the results fall.
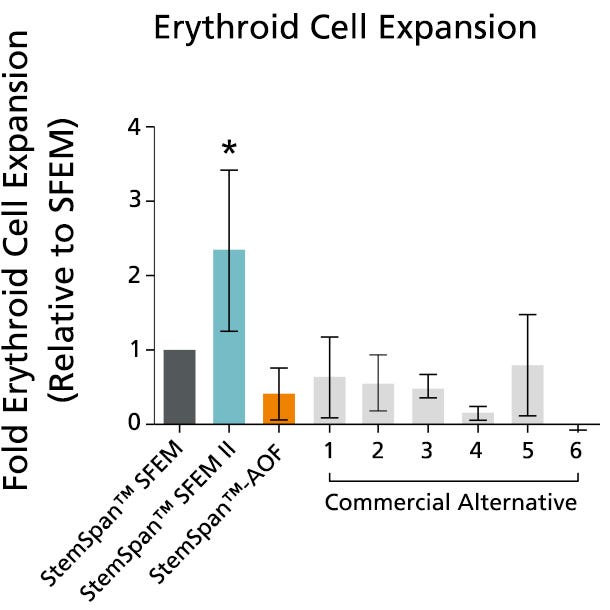
Figure 5. StemSpan™ SFEM II Serum-Free Expansion Medium Containing Erythroid Expansion Supplement Supports Greater Expansion of Erythroid Cells Than Other Media Tested
The numbers of erythroid cells, normalized relative to the values obtained in StemSpan™ SFEM medium (dark gray bar), obtained after culturing purified CD34+ CB cells for 14 days in StemSpan™ SFEM, SFEM II (blue bar) and AOF (orange bar), and six media from other commercial suppliers (light gray bars, commercial alternative 1-6, which included, in random order, X-Vivo-15 and HPGM (both from Lonza), StemLine II (Sigma), HP01 (Macopharma), StemPro34 (Life Technologies) and SCGM (Cellgenix). All media were supplemented with StemSpan™ Erythroid Expansion Supplement (Catalog #02692). Vertical lines indicate 95% confidence limits, the range within which 95% of results fall. The numbers of cells produced in StemSpan™ SFEM II were significantly higher than in all other media (*p<0.05, paired t-test, n=6).
Note: Data for StemSpan™-AOF shown were generated with the original phenol red-containing version StemSpan™-ACF (Catalog #09855). However internal testing showed that the performance of the new phenol red-free, cGMP-manufactured version, StemSpan™-AOF (Catalog #100-0130) was comparable.
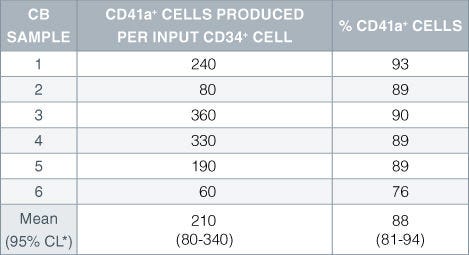
Table 2. Production of Megakaryocytes From CD34+ Human Cord Blood Cells Cultured in StemSpan™ SFEM Serum-Free Expansion Medium Containing Megakaryocyte Expansion Supplement
Numbers and percent of cells expressing the megakaryocyte marker CD41a produced after 14 days of culture of enriched CD34 + cells from 6 independent cord blood (CB) samples. *95% confidence limits, the range within which 95% of the results fall.
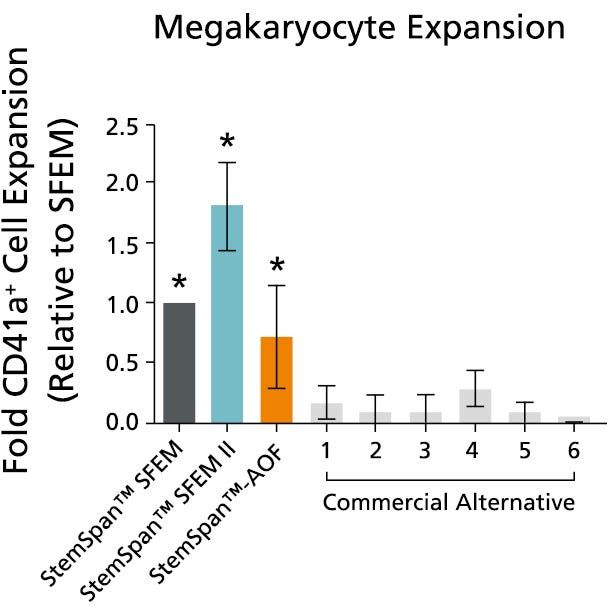
Figure 6. StemSpan™ SFEM II Serum-Free Expansion Medium Containing Megakaryocyte Expansion Supplement Supports Greater Expansion of Megakaryocytes Than Other Media Tested
The numbers of megakaryocytes, normalized relative to the values obtained in StemSpan™ SFEM medium (dark gray bar), obtained after culturing purified CD34+ CB cells for 14 days in StemSpan™ SFEM, SFEM II (blue bar) and AOF (orange bar), and six media from other commercial suppliers (light gray bars, Commercial Alternative 1-6, which included, in random order, StemLine II (Sigma), HPGM (Lonza), HP01 (Macopharma), SCGM (Cellgenix), StemPro34 (Life Technologies) and X-Vivo-15 (Lonza). All media were supplemented with StemSpan™ Megakaryocyte Expansion Supplement (Catalog #02696). Vertical lines indicate 95% confidence limits, the range within which 95% of results fall. The numbers of cells produced in the StemSpan™ media were significantly higher than in the other media (*p<0.01 paired t-test, n=6).
Note: Data for StemSpan™-AOF shown were generated with the original phenol red-containing version StemSpan™-ACF (Catalog #09855). However internal testing showed that the performance of the new phenol red-free, cGMP-manufactured version, StemSpan™-AOF (Catalog #100-0130) was comparable.
This product is designed for use in the following research area(s) as part
of the highlighted workflow stage(s). Explore these workflows to learn more about the other products we
offer to support each research area.
Enhancement of proliferation of human umbilical cord blood-derived CD34+ hematopoietic stem cells by a combination of hyper-interleukin-6 and small molecules.
Y. S. Park et al.
Biochemistry and biophysics reports 2022 mar
Abstract
Umbilical cord blood (UCB) is an alternative source of allogeneic hematopoietic stem cells (HSCs) for transplantation to treat various hematological disorders. The major limitation to the use of UCB-derived HSCs (UCB-HSCs) in transplantation, however, is the low numbers of HSCs in a unit of cord blood. To overcome this limitation, various cytokines or small molecules have been used to expand UCB-HSCs ex vivo. In this study, we investigated a synergistic effect of the combination of HIL-6, SR1, and UM171 on UCB-HSC culture and found that this combination resulted in the highest number of CD34+ cells. These results suggest that the combination of SR1, UM171 and HIL-6 exerts a synergistic effect in the proliferation of HSCs from UCB and thus, SR1, UM171 and HIL-6 is the most suitable combination for obtaining HSCs from UCB for clinical transplantation.
Protocol to identify and analyze mouse and human quiescent hematopoietic stem cells using flow cytometry combined with confocal imaging.
J. Qiu et al.
STAR protocols 2022 dec
Abstract
Mitochondrial membrane potential (MMP) segregates functionally distinct subsets within highly purified hematopoietic stem cells (HSCs). Here, we detail a protocol for FACS isolation of MMP sub-fractions of phenotypically defined mouse and human HSCs. These steps are followed by high-/super-resolution immunofluorescence microscopy of HSCs' lysosomes. While the protocol describes the isolation of quiescent HSCs, which are the most potent subsets, it could also be applied to other HSC subsets. This protocol overcomes some experimental challenges associated with low HSC numbers. For complete details on the use and execution of this protocol, please refer to Liang et al. (2020) and Qiu et al. (2021).
Reducing TGF-$\beta$1 cooperated with StemRegenin 1 promoted the expansion ex vivo of cord blood CD34+ cells by inhibiting AhR signalling.
X. Zhu et al.
Cell proliferation 2021 mar
Abstract
OBJECTIVE As an inhibitor of the AhR signalling pathway, StemRegenin 1 (SR1) not only promotes the expansion of CD34+ cells but also increases CD34- cell numbers. These CD34- cells influenced the ex vivo expansion of CD34+ cells. In this work, the effects of periodically removing CD34- cells combined with SR1 addition on the ex vivo expansion and biological functions of HSCs were investigated. MATERIALS AND METHODS CD34- cells were removed periodically with SR1 addition to investigate cell subpopulations, cell expansion, biological functions, expanded cell division mode and supernatant TGF-$\beta$1 contents. RESULTS After 10-day culture, the expansion of CD34+ cells in the CD34- cell removal plus SR1 group was significantly higher than that in the control group and the SR1 group. Moreover, periodically removing CD34- cells with SR1 addition improved the biological function of expanded CD34+ cells and significantly increased the percentage of self-renewal symmetric division of CD34+ cells. In addition, the concentration of total TGF-$\beta$1 and activated TGF-$\beta$1 in the supernatant was significantly lower than those in the control group and the SR1 group. RT-qPCR results showed that the periodic removal of CD34- cells with cooperation from SR1 further reduced the expression of AhR-related genes. CONCLUSIONS Periodic removal of CD34- cells plus cooperation with SR1 improved the expansion of CD34+ cells, maintained better biological function of expanded CD34+ cells and reduced the TGF-$\beta$1 contents by downregulating AhR signalling.
PRODUCTS ARE FOR RESEARCH USE ONLY AND NOT INTENDED FOR HUMAN OR ANIMAL DIAGNOSTIC OR THERAPEUTIC USES UNLESS OTHERWISE STATED. FOR ADDITIONAL INFORMATION ON QUALITY AT STEMCELL, REFER TO WWW.STEMCELL.COM/COMPLIANCE.