Thank you for your interest in this product.
Please provide us with your contact information and your local representative
will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration
By submitting this form, you are providing your consent to STEMCELL Technologies Canada Inc. and its subsidiaries and affiliates (“STEMCELL”) to collect and use your information, and send you newsletters and emails in accordance with our privacy policy. Please contact us with any questions that you may have. You can unsubscribe or change your email preferences at any time.
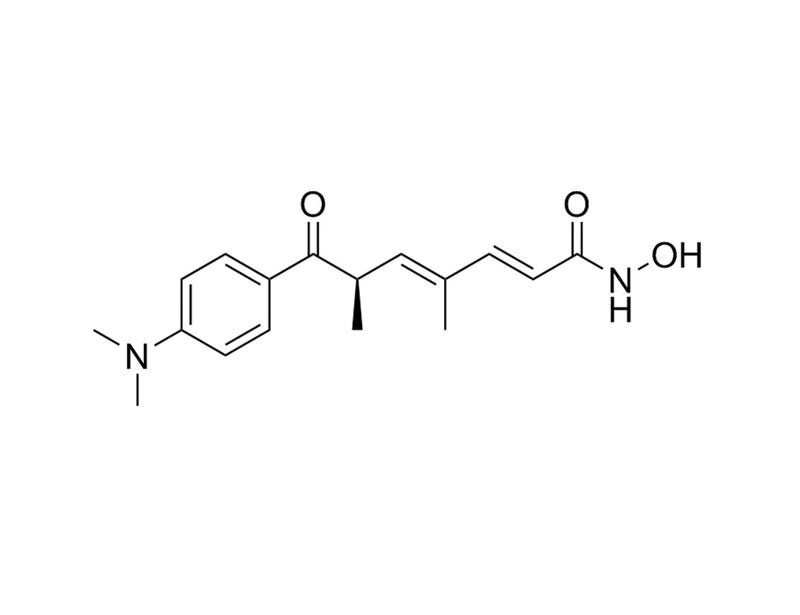
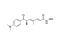
Trichostatin A is a potent and reversible inhibitor of Histone Deacetylase (HDAC), therefore acting as an epigenetic modifier by preventing the removal of acetyl groups from lysine residues on histone tails. Trichostatin A inhibits both class I and class II HDACs, including HDAC1 (IC₅₀ = 6 nM), HDAC4 (IC₅₀ = 38 nM), and HDAC6 (IC₅₀ = 8.6 nM). (Furumai et al.; Yoshida et al.)
REPROGRAMMING
· Increases the reprogramming efficiency of mouse embryonic fibroblasts to induced pluripotent stem (iPS) cells (Huangfu et al.).
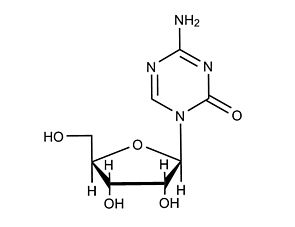
· Resets epigenetic memory in mouse iPS cells, in combination with 5-Azacytidine (Kim et al.).
·Increases the efficiency of cloned mouse embryo development by somatic cell nuclear transfer (Kishigami et al.).
MAINTENANCE AND SELF-RENEWAL
· Prevents dedifferentiation of primary rat hepatocytes in culture, maintaining liver-specific cellular functions (Henkens et al.).
DIFFERENTIATION
· Promotes differentiation of hepatocytes from human mesenchymal stem cells (Snykers et al.).
Cell Type
Hepatic Cells, Mesenchymal Stem and Progenitor Cells, Pluripotent Stem Cells
This product is designed for use in the following research area(s) as part
of the highlighted workflow stage(s). Explore these workflows to learn more about the other products we
offer to support each research area.
Epigenetic memory in induced pluripotent stem cells.
Kim K et al.
Nature 2010 SEP
Abstract
Somatic cell nuclear transfer and transcription-factor-based reprogramming revert adult cells to an embryonic state, and yield pluripotent stem cells that can generate all tissues. Through different mechanisms and kinetics, these two reprogramming methods reset genomic methylation, an epigenetic modification of DNA that influences gene expression, leading us to hypothesize that the resulting pluripotent stem cells might have different properties. Here we observe that low-passage induced pluripotent stem cells (iPSCs) derived by factor-based reprogramming of adult murine tissues harbour residual DNA methylation signatures characteristic of their somatic tissue of origin, which favours their differentiation along lineages related to the donor cell, while restricting alternative cell fates. Such an 'epigenetic memory' of the donor tissue could be reset by differentiation and serial reprogramming, or by treatment of iPSCs with chromatin-modifying drugs. In contrast, the differentiation and methylation of nuclear-transfer-derived pluripotent stem cells were more similar to classical embryonic stem cells than were iPSCs. Our data indicate that nuclear transfer is more effective at establishing the ground state of pluripotency than factor-based reprogramming, which can leave an epigenetic memory of the tissue of origin that may influence efforts at directed differentiation for applications in disease modelling or treatment.
Induction of pluripotent stem cells by defined factors is greatly improved by small-molecule compounds
Huangfu D et al.
Nat Biotechnol 2008
Abstract
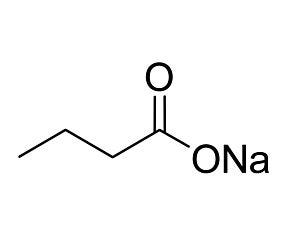
Reprogramming of mouse and human somatic cells can be achieved by ectopic expression of transcription factors, but with low efficiencies. We report that DNA methyltransferase and histone deacetylase (HDAC) inhibitors improve reprogramming efficiency. In particular, valproic acid (VPA), an HDAC inhibitor, improves reprogramming efficiency by more than 100-fold, using Oct4-GFP as a reporter. VPA also enables efficient induction of pluripotent stem cells without introduction of the oncogene c-Myc.
Chromatin remodeling agent trichostatin A: a key-factor in the hepatic differentiation of human mesenchymal stem cells derived of adult bone marrow.
Snykers S et al.
BMC developmental biology 2007 JAN
Abstract
BACKGROUND The capability of human mesenchymal stem cells (hMSC) derived of adult bone marrow to undergo in vitro hepatic differentiation was investigated. RESULTS Exposure of hMSC to a cocktail of hepatogenic factors [(fibroblast growth factor-4 (FGF-4), hepatocyte growth factor (HGF), insulin-transferrin-sodium-selenite (ITS) and dexamethasone)] failed to induce hepatic differentiation. Sequential exposure to these factors (FGF-4, followed by HGF, followed by HGF+ITS+dexamethasone), however, resembling the order of secretion during liver embryogenesis, induced both glycogen-storage and cytokeratin (CK)18 expression. Additional exposure of the cells to trichostatin A (TSA) considerably improved endodermal differentiation, as evidenced by acquisition of an epithelial morphology, chronological expression of hepatic proteins, including hepatocyte-nuclear factor (HNF)-3beta, alpha-fetoprotein (AFP), CK18, albumin (ALB), HNF1alpha, multidrug resistance-associated protein (MRP)2 and CCAAT-enhancer binding protein (C/EBP)alpha, and functional maturation, i.e. upregulated ALB secretion, urea production and inducible cytochrome P450 (CYP)-dependent activity. CONCLUSION hMSC are able to undergo mesenchymal-to-epithelial transition. TSA is hereby essential to promote differentiation of hMSC towards functional hepatocyte-like cells.
PRODUCTS ARE FOR RESEARCH USE ONLY AND NOT INTENDED FOR HUMAN OR ANIMAL DIAGNOSTIC OR THERAPEUTIC USES UNLESS OTHERWISE STATED. FOR ADDITIONAL INFORMATION ON QUALITY AT STEMCELL, REFER TO WWW.STEMCELL.COM/COMPLIANCE.