Make more informed purchasing decisions with our new product availability and delivery estimate feature, now available on all product pages, in your cart, and during checkout.
Sign In
New to STEMCELL?
Register for an account to quickly and easily purchase products online and for one-click access to all educational content.
Thank you for your interest in this product.
Please provide us with your contact information and your local representative
will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration
By submitting this form, you are providing your consent to STEMCELL Technologies Canada Inc. and its subsidiaries and affiliates (“STEMCELL”) to collect and use your information, and send you newsletters and emails in accordance with our privacy policy. Please contact us with any questions that you may have. You can unsubscribe or change your email preferences at any time.
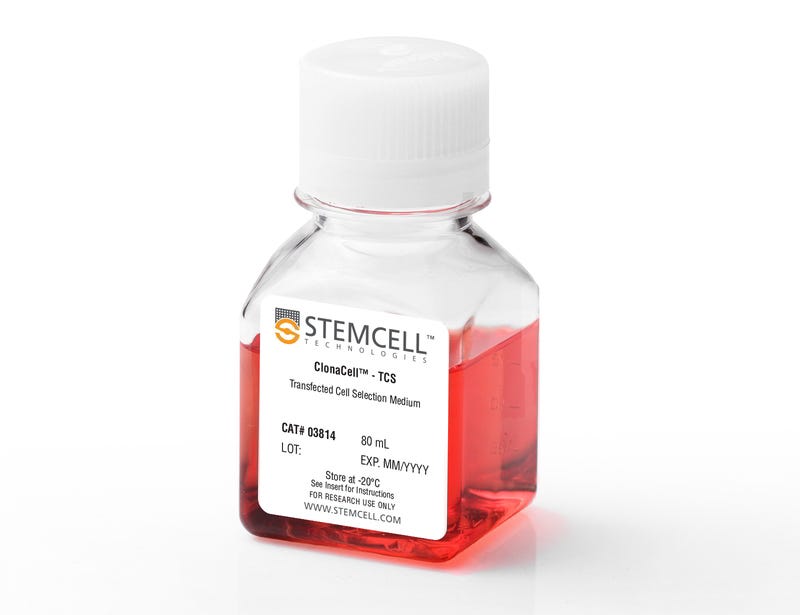
ClonaCell™-TCS Medium is a serum-containing methylcellulose-based semi-solid medium that can be used for selecting and cloning a variety of suspension-adapted cell lines, including CHO-S and hybridomas. It can also be used for semi-solid cloning of some cell lines that grow adherently in the presence of serum, including CHO-K1, BHK-1, and HEK-293. This medium contains pre-selected fetal bovine serum (FBS) and bovine serum albumin (BSA) and supports robust growth of a wide variety of cell types. ClonaCell™-TCS Medium does not contain selection agents.
Benefits of semi-solid cloning:
• Individual cells are suspended in viscous medium and form physically separated, discrete colonies that are easily isolated.
• Monoclonal cell lines are isolated in less time using fewer resources compared with selection and cloning by limiting dilution.
• Diverse clones with a wide range of growth rates and productivities form discrete colonies in the viscous medium. As a result, rare and high-producing clones can be individually isolated more easily using simultaneous selection and cloning in semi-solid medium compared with selection in bulk liquid cultures.
Contains
• IMDM (Iscove's Modified Dulbecco's Medium)
• Methylcellulose
• Pre-selected serum
• Bovine serum albumin
• 2-Mercaptoethanol
• Phenol red
• L-Glutamine
• Other ingredients
Subtype
Semi-Solid Media, Specialized Media
Cell Type
CHO Cells, HEK-293, Hybridomas, Other
Species
Mouse, Other
Application
Cell Culture, Semi-Solid Cloning
Brand
ClonaCell
Area of Interest
Antibody Development, Cell Line Development, Hybridoma Generation
This product is designed for use in the following research area(s) as part
of the highlighted workflow stage(s). Explore these workflows to learn more about the other products we
offer to support each research area.
Is the serum in ClonaCell™-TCS medium heat inactivated?
Yes, all serum used in ClonaCell™ is heat inactivated.
Is there any IgG in ClonaCell™ TCS?
While we don't add IgG to the ClonaCell™ media, we do add serum, which contains an undefined amount of IgG. We selectively use serum lots with low IgG levels in the production of ClonaCell™ media, however, levels vary from lot to lot. IgG levels in a specific lot of ClonaCell™ TCS medium are available in the lot-specific Certificate of Analysis.
Can ClonaCell™-TCS be used with any cell line?
A list of recommended cell lines can be found in the manual. Other cell lines may be compatible with ClonaCell™-TCS. It will be necessary, however, to determine the plating cell density and growth efficiency of the desired cells in ClonaCell™-TCS.
Why do I get more cells when I select my fusion in liquid medium rather than in methylcellulose-based semi-solid medium?
Cells grown in liquid medium may appear to grow more rapidly than in methylcellulose-based medium. This is often due to the presence of a few rapidly growing clones that multiply quickly and become abundant in liquid culture, overgrowing clones that grow more slowly. In methylcellulose cultures, the rapidly growing cells remain in close proximity to each other, resulting in large colonies, each derived from a single fusion or transfection product. The large clones don't overgrow smaller, slower growing colonies, which can be separately isolated.
How do I thaw ClonaCell™ methylcellulose-based semi-solid medium?
We recommend thawing the medium overnight in a refrigerator at 4°C and mixing well.
How do I measure and dispense methylcellulose semi-solid medium?
We recommend using a 12 mL syringe with a 16 gauge needle attached (blunt end needles are recommended for safety purposes). Do not dispense the semi-solid media/cell mixture using serological pipettes as the media will stick to the pipette walls, resulting in inaccurate dispensed volumes and loss of cells.
My ClonaCell™ methylcellulose semi-solid medium appears runny. Why does this happen?
"Runny" methylcellulose could be a result of improper handling. Diluting the methylcellulose with too much liquid medium, or insufficient mixing before use, will result in methylcellulose with altered viscosity. Excessive condensation on the inside of the cell culture dish lid can result in water dripping onto the cultures, lowering viscosity. Additionally, bumping, shaking or other sudden movement of the culture may also disrupt the colonies. Note: methylcellulose is less viscous at room temperature than at 37°C.
What is the optimal number of colonies per plate?
We recommend 50-150 colonies per plate. As it is difficult to anticipate the numbers of colonies after a fusion or transfection, we recommend plating at three different densities to increase the likelihood of achieving a plating density of approximately 100 colonies per plate. This density allows sufficient space between the colonies to allow for easy colony picking.
There are still bubbles in the media after I plate my cells. Do I need to disrupt the bubbles?
We recommend that you avoid creating large bubbles during plating, but there is no need to manually pop or disperse the small bubbles after plating. They will disperse over the incubation period of 10-14 days.
Do I ever need to re-clone cultures grown with ClonaCell™ semi-solid medium?
Re-cloning is a good practice to observe and is recommended if the number of colonies in the original dishes was very high.
Once I pick the colonies and grow the cells in plates, will the residual methylcellulose interfere with characterization? For example, will I have problems doing an ELISA?
There will likely be some residual methylcellulose contamination when colonies are picked and transferred to the 96-well plate with the liquid growth medium. The concentration of methylcellulose, however, should be low enough that it should not interfere with most assays.
How important is the incubator humidity when culturing in methylcellulose-based medium?
Very important. In situations where the humidity is not high enough, we recommend that the 100 mm Petri dishes should be placed with an open dish containing sterile water inside a larger plastic container with a lid. Without very high humidity, the media will dry out over the culture period and this will impede the growth of the colonies.
Do I have to use 100 mm petri dishes or can I use other cultureware?
We recommend 100 mm Petri dishes as these have been used to develop and test ClonaCell™ semi-solid media. We have found that the surface area of these dishes allows for easy colony picking. Other sizes of dish (e.g. 6-well plates) can be used. It is important to use non-coated dishes to prevent cells from sticking to the bottom of the plate and obscuring the colonies. The volume of media plated should be adjusted to reflect the surface area of the dish being used.
The T-cell leukemia-associated ribosomal RPL10 R98S mutation enhances JAK-STAT signaling.
T. Girardi et al.
Leukemia 2018 MAR
Abstract
Several somatic ribosome defects have recently been discovered in cancer, yet their oncogenic mechanisms remain poorly understood. Here we investigated the pathogenic role of the recurrent R98S mutation in ribosomal protein L10 (RPL10 R98S) found in T-cell acute lymphoblastic leukemia (T-ALL). The JAK-STAT signaling pathway is a critical controller of cellular proliferation and survival. A proteome screen revealed overexpression of several Jak-Stat signaling proteins in engineered RPL10 R98S mouse lymphoid cells, which we confirmed in hematopoietic cells from transgenic Rpl10 R98S mice and T-ALL xenograft samples. RPL10 R98S expressing cells displayed JAK-STAT pathway hyper-activation upon cytokine stimulation, as well as increased sensitivity to clinically used JAK-STAT inhibitors like pimozide. A mutually exclusive mutation pattern between RPL10 R98S and JAK-STAT mutations in T-ALL patients further suggests that RPL10 R98S functionally mimics JAK-STAT activation. Mechanistically, besides transcriptional changes, RPL10 R98S caused reduction of apparent programmed ribosomal frameshifting at several ribosomal frameshift signals in mouse and human Jak-Stat genes, as well as decreased Jak1 degradation. Of further medical interest, RPL10 R98S cells showed reduced proteasome activity and enhanced sensitivity to clinical proteasome inhibitors. Collectively, we describe modulation of the JAK-STAT cascade as a novel cancer-promoting activity of a ribosomal mutation, and expand the relevance of this cascade in leukemia.
DNMT3A mutations promote anthracycline resistance in acute myeloid leukemia via impaired nucleosome remodeling.
Guryanova OA et al.
Nature Medicine 2016 NOV
Abstract
Although the majority of patients with acute myeloid leukemia (AML) initially respond to chemotherapy, many of them subsequently relapse, and the mechanistic basis for AML persistence following chemotherapy has not been determined. Recurrent somatic mutations in DNA methyltransferase 3A (DNMT3A), most frequently at arginine 882 (DNMT3A(R882)), have been observed in AML and in individuals with clonal hematopoiesis in the absence of leukemic transformation. Patients with DNMT3A(R882) AML have an inferior outcome when treated with standard-dose daunorubicin-based induction chemotherapy, suggesting that DNMT3A(R882) cells persist and drive relapse. We found that Dnmt3a mutations induced hematopoietic stem cell expansion, cooperated with mutations in the FMS-like tyrosine kinase 3 gene (Flt3(ITD)) and the nucleophosmin gene (Npm1(c)) to induce AML in vivo, and promoted resistance to anthracycline chemotherapy. In patients with AML, the presence of DNMT3A(R882) mutations predicts minimal residual disease, underscoring their role in AML chemoresistance. DNMT3A(R882) cells showed impaired nucleosome eviction and chromatin remodeling in response to anthracycline treatment, which resulted from attenuated recruitment of histone chaperone SPT-16 following anthracycline exposure. This defect led to an inability to sense and repair DNA torsional stress, which resulted in increased mutagenesis. Our findings identify a crucial role for DNMT3A(R882) mutations in driving AML chemoresistance and highlight the importance of chromatin remodeling in response to cytotoxic chemotherapy.
Kit containing ClonaCell™-TCS Medium and hygromycin B antibiotic
Item added to your cart
ClonaCell™-TCS Medium
Quality Statement:
PRODUCTS ARE FOR RESEARCH USE ONLY AND NOT INTENDED FOR HUMAN OR ANIMAL DIAGNOSTIC OR THERAPEUTIC USES UNLESS OTHERWISE STATED. FOR ADDITIONAL INFORMATION ON QUALITY AT STEMCELL, REFER TO WWW.STEMCELL.COM/COMPLIANCE.