Airway Epithelial Cells of the Human Respiratory System
Learn about the airway epithelium of the human respiratory system and in vitro human airway modeling
Request Pricing
Thank you for your interest in this product. Please provide us with your contact information and your local representative will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration

- Document # DX20359
- Version 1.0.0
- Apr 2015
Airway Epithelium and Its Region- Specific Stem And Progenitor Cells
Human Airway Epithelium
The human respiratory system functions to transfer oxygen from the air to the circulatory system and carbon dioxide from the body to the surrounding air. The airway warms and moistens the air prior to gas exchange as it travels towards the distal lung, and protects the distal lung from harmful pathogens and environmental insults. The human airway can be divided into the conducting zone and the respiratory zone. The conducting zone includes regions that do not participate in gas exchange, namely the nasal passage, trachea, main bronchi, intra-pulmonary bronchi and bronchioles. The diameter and length of each successive airway branch decreases progressively from the trachea to the terminal bronchioles. The respiratory zone consists of the respiratory bronchioles, alveolar ducts and alveolar sacs. The respiratory zone is the site of gas exchange between the air within the lung and the blood within pulmonary capillaries.1,2
The human airway, from the nasal passage to the alveolar sacs, is covered with a continuous epithelial sheet that differs in morphology and cellular composition between the conducting and respiratory zones. In the most proximal regions of the conducting zone, including the nasal passage, trachea, and bronchi, the airway epithelium exhibits a columnar, pseudostratified morphology. The height of this epithelium decreases in more distal regions of the conducting zone and resembles a cuboidal epithelium in the small airway. The major cell types of the large airway epithelium are goblet cells that produce and secrete mucus, ciliated cells that promote mucus motility through coordinated movement of their apical cilia, and basal cells that line the basement membrane and do not contact the apical surface of the epithelium. In the bronchioles, the cuboidal epithelium contains secretory club cells and fewer ciliated cells than in more proximal airway regions. The alveolar epithelium is lined with type I and II alveolar epithelial cells (AECs). The alveolar cells fuse to endothelial cells by their basal membranes to form the gas exchange barrier. There is a variety of less frequent cell types throughout the airway, including neuroendocrine cells in the large airway, and a resident population of alveolar macrophages (phagocytes) that reside in the interstitial spaces and function to remove inhaled debris deposited in the air spaces.1,3
Epithelial Barriers and Defense Function
The airway epithelium plays a critical role in conducting air to and from the alveoli. It is central to the defense of the lung against pathogens and particulates that are inhaled from the environment. The primary action of defense is the combined function of secretory and ciliated cells maintaining an efficient mucociliary elevator that promotes expulsion of foreign material.4,5 A variety of other host defense processes also contribute to the efficiency of the protective function of the airway epithelium. Importantly, the epithelial sheet does not function as an independent entity, but rather as an interdependent functional unit along with other epithelial cells, mesenchymal cells, endothelial cells and the extracellular matrix.6,7,8
An additional layer of defense is achieved by intercellular junctions that create an efficient barrier against inhaled pathogens or toxic agents. The permeability barrier is regulated by tight junctions that limit the passive flow of molecules between the apical and basolateral compartments. Tight junctions are also key regulators of ion and solute homeostasis, thus effecting the control over the height and ionic composition of the airway surface liquid that is necessary for lung defense.9
Despite these efficient defense systems, the airway epithelium is in permanent contact with the external environment and is frequently injured. After airway epithelial injury, the basement membrane may be partially or completely denuded, with only clusters of basal cells remaining attached to the basement membrane. These typical alterations in structure and function play a major role in the pathogenesis of numerous respiratory diseases, such as chronic bronchitis, asthma, chronic obstructive pulmonary disease (COPD) or cystic fibrosis (CF), and can originate from exposure to everyday pulmonary inhalants such as natural and man-made particulates, fibrous materials and inhaled chemicals. Immediately after injury, the airway epithelium initiates a repair process to restore barrier integrity and normal epithelial function, but these processes are not fully understood.1,5,10,11
Airway Region-specific Stem and Progenitor Cells
The airway epithelium is a dynamic tissue that normally undergoes slow but constant turnover. In the event of mild to moderate injury, the airway epithelium responds vigorously to re-establish an epithelial sheet with normal structure and function. This repair process is carried out by the airway stem and progenitor cells.11-18
The vocabulary describing the different progenitor cells is complex in the field of epithelial progenitor cell biology. In this review, we use progenitor to broadly define any relatively undifferentiated cell that is capable of proliferation and differentiation, while stem cells are a rare subtype of progenitor cells that are capable of sustained proliferation, self-renewal and multipotent differentiation. In contrast, many adult progenitor cells are transient amplifying cells, which proliferate extensively to establish terminally differentiated cells, but have a limited life span in comparison to stem cells.12,13,19
Evidence suggests that different kinds of epithelial stem and progenitor cells reside in different regions of the lung: basal cells in the proximal trachea and bronchi,20 bronchioalveolar stem cells (BASCs) in the bronchial-alveolar duct junction (BADJ),21 and alveolar type 2 epithelial cells in the alveolar region.22
Basal Cells
There is now increasing evidence that basal cells are multipotent progenitor cells in the tracheobronchial region that can both self-renew and give rise to ciliated and secretory lineages during postnatal growth and steady-state periods, as well as during repair following damage to the epithelium. This understanding is derived from in vivo genetic lineage tracing studies and in vitro culture including air-liquid interface (ALI) culture and 3D clonal sphere culture. Using both of these culture techniques, the potential of single cells to self-renew and differentiate can be followed.20,23,24 Either of these approaches rely on the ability to accurately tag basal cells using markers whose functional importance to the biology of the basal cell progenitor remains unclear. The most widely used basal cell markers include P63, IGTA6, NGFR, KRT5 and KRT14.19,20,23-27 Of these markers, KRT5 and KRT14 are most commonly used. While KRT5 is expressed in all basal cells, only a subset of basal cells normally expresses KRT14 in steady-state when the turnover of the epithelium is low.23 This suggests that there may be functional heterogeneity among individual basal cells, with only some being able to give rise to multiple mature cell lineages. Lineage tracing studies in mouse tracheal and bronchial epithelium using a KRT14-CreER driver suggest that subpopulations of basal cells may have multipotent or unipotent capacities following airway injury. The KRT5 and KRT14 double-positive basal cells that are present singly or in small clusters transiently increase during repair following damage to the luminal cells by a variety of agents such as acid, detergent, or systemic naphthalene administration. This result suggests that KRT14 is up-regulated in basal cells when they are ‘activated’ to proliferate.23,28 Other studies, however, have proposed that all basal cells have the capacity for multipotent differentiation and that the fate of the basal cell progenitor is influenced by its local environment and by the mechanism of injury.20
One question of practical relevance for regenerative therapies in the human lung is whether basal cells are the only cells that can efficiently repair the pseudostratified epithelium or whether differentiated cells can dedifferentiate or transdifferentiate under certain conditions. A key paper published recently from the Rajagopal lab used a strategy to specifically kill KRT5+ basal cells in the mouse trachea in vivo.29 Under these conditions they found that differentiated SCGB1A1+ secretory cells can undergo dedifferentiation into P63 and KRT5 double-positive basal cells. These basal cells persist long-term and behave like normal KRT5+ progenitors. The authors further demonstrate through in vitro cultures that contact with basal cells prevents luminal cell dedifferentiation, but the precise mechanisms driving reprogramming and subsequent stem cell function need further study.29
Bronchioalveolar Stem Cells (BASCs)
The transition zone between the smallest terminal bronchioles and the alveoli is known as the BADJ. In the mouse, this region contains both ciliated and secretory (club) cells, but in humans the resident cell type(s) are poorly defined. In mice, a small number of cells in the BADJ (about 1–2 per duct) co-express Scgb1a1 (secretory cells) and surfactant protein C (Sftpc), a protein that is expressed at highest levels by Type II AECs cells in the alveoli. It has been proposed that these double-positive cells are BASCs. This designation is largely based on the fact that when isolated by FACS, the cells can give rise to bronchiolar and alveolar cell types in culture.30 Kim and her colleagues further identified a BMP4-controlled NFATC1-TSP axis in lung endothelial cells that directs BASCs to differentiate into the alveolar lineage using a state-of-the-art 3D clonal co-culture system.21
Type II Alveolar Epithelial Cells (AEC2s)
The major epithelial cell types of the gas exchanging region are cuboidal AEC2s that are specialized for surfactant protein production and secretion, and flat AEC1s that are specialized for gas exchange. Cell turnover in the alveolar region of the adult mouse lung is normally slow, making it difficult to follow steady-state lineage relationships. After experimental injuries that damage alveolar cells and/or expose them to oxidative stress, however, there is rapid proliferation of surviving cells and activation of repair mechanisms.
Studies now more than 40 years old that used H3-thymidine labeling showed that AEC2s in adult monkeys and rats proliferate in response to injury by hyperoxia and nitric oxide and give rise to AEC1s. This capacity for self-renewal and differentiation of adult AEC2s has been confirmed by recent in vivo genetic lineage tracing studies using Cre recombinase driven by genes associated with functions of the terminally differentiated cells including Sftpc and Lyz2. During steady state turnover, there is relatively little clonal expansion of individual AEC2s and very little differentiation into AEC1s. After injury of the alveolar region by bleomycin and by hyperoxia, the rate of differentiation of lineage labeled AEC2s into AEC1s is much higher.22,31-34
The identification of stem and progenitor lineages and activity in the adult has proceeded much faster than our understanding of the molecular pathways that regulate their cellular behavior. The concept that developmental pathways are reactivated and play important roles in lung repair and regeneration requires additional testing, especially in more physiologically or clinically relevant models. These studies will require additional information on how structures are formed in the lung, in particular the process of alveologenesis, which is still poorly understood. This is a topic that will greatly benefit from additional research into the development and maturation of the lung. Importantly, the advent of new in vitro models of lung stem cell activity such as tracheospheres and alveolospheres should allow for testing of factors that can promote either stem/progenitor self-renewal or differentiation at the single-cell level.
In Vitro Human Airway Modeling
To appropriately model the in vivo human airway, specialized in vitro cell culture techniques have been developed that promote the formation of 3D structures that recapitulate the morphological and functional characteristics of the airway. For example, basal cells from the proximal human airway, upon recapitulation, differentiate into a mucociliary pseudostratified epithelium containing ciliated, goblet and basal cells. In this in vitro cell culture environment, cells display beating cilia, mucus secretion, barrier properties and remodeling and restoration properties similar to the native airway epithelium.1,35
Two dominant tissue-engineering techniques have been developed to model the native lung environment in vitro. Models are available for all of the major regions of the respiratory tract,36 however, clinical and basic science applications have focused primarily on recapitulation of the bronchial epithelium. This is likely due to the fact that numerous pathologies result from disruption of this region and that researchers can obtain primary cell samples from this region with relative ease.
The most well-characterized of these models is the 3D organotypic ALI culture on tissue culture inserts.36-43 In this method, the human bronchial epithelial cells obtained from bronchial brushings or through isolation of epithelial cells from cadaveric donor tissue can be seeded on permeable inserts. The procedure for inducing cell differentiation first requires growth of the cells in submerged cultures on inserts until they become fully confluent, after which they are raised to the ALI and cultured until they have become fully differentiated, a process that takes between 21 and 28 days. Once ‘air-lifted’, ALI cultures are fed with medium bathing the basolateral cell surface only. The combination of apical exposure to air and basal exposure to specialized cell culture medium triggers differentiation of the monolayer culture to a pseudostratified columnar epithelium that resembles the native human airway. This model system is useful in a number of applications ranging from in vivo inhalation toxicology and drug screening to basic research.44 The advantages of such a model over in vivo animal studies include the use of human tissue, a relatively low cost of maintenance and ease of handling. The use of ALI cultures also offers a unique opportunity for the direct deposition of aerosol particles onto semi-dry apical cell surfaces, a procedure that mimics the deposition of powders onto the airway surface.1,45,46
Isolated airway epithelial cells can also be cultured as 3D spheres in semi-solid Matrigel® Matrix with or without the use of tissue culture inserts. In this model, individual cells are able to grow into clonal spheres called bronchospheres or alveolospheres, depending on the origin of the starting cells.20-22 While bronchospheres can be formed in specialized matrices and medium without co-culture with other tissues, using current methods, alveolospheres can only be formed by co-culturing with other tissue types, such as fibroblasts or endothelial cells.21,23 The spheres consist of a lumen surrounded by an epithelial cell layer, with the apical surface of Goblet and ciliated cells pointing into the lumen. Following the proper procedure, the sphere culture can be achieved without tissue culture inserts, making it an attractive candidate system for high-throughput screening.
Find the Right Media for Your Pulmonary Research
Whether you’re starting with primary airway epithelial cells or human pluripotent stem cells, finding the right media can help you generate reproducible cultures and standardize your pulmonary workflows. Use our Interactive Product Finder to generate a complete list of materials to support your pulmonary workflow, whether you’re culturing your cells as monolayers, organoids, or at the air-liquid interface (ALI).
Future Outlook
Modeling of airway epithelial cells using organotypic culture systems is gaining significant popularity with the basic research community, as well as with the pharmaceutical industry and contract research organizations performing disease modeling and efficacy assays as part of upstream drug development processes. Adoption of these techniques is driven primarily by the motivation to move to a more physiologically representative model system than the animal studies and immortalized cell lines that previously represented the best options for investigating airway biology. The ability to culture human primary cells as differentiated epithelial layers has increased both the relevance and scope of experiments that can be carried out in vitro, compared to previous techniques. Using either ALI or matrix-suspended sphere culture techniques, researchers can now interrogate the functional properties of differentiated human airway cells with respect to specific experimental variables in vitro. Additionally, the ability to culture primary human cells in these organotypic systems allows for experiments incorporating the genotype of specific patients, opening the door for individualized diagnostic and treatment options.
Featured Content and Resources
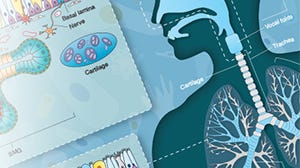
Respiratory System Overview
Get a free wallchart providing an organizational and functional overview of the respiratory system.

Optimizing In Vitro Lung Models
Dr. Rachael Rayner compares the effects of various expansion and differentiation media on primary airway cells cultured at the ALI.
References
- Berube K, et al. Altern Lab Anim 37: 89-141, 2009
- Crystal RG, et al Proc Am Thorac Soc 5: 772-777, 2008
- Kaisani A, et al. Differentiation 87: 119-126, 2014
- Randell SH. Proc Am Thorac Soc 3: 718-725, 2006
- Livraghi A and Randell SH. Toxicol Pathol 35: 116-129, 2007
- Rock JR, et al. Proc Natl Acad Sci U S A 108: E1475-1483, 2011
- Weaver M, et al. Dev Biol 258: 169-184, 2003
- Knight DA, et al. Expert Rev Respir Med. 4(6): 747-58, 2010
- Matsui H, et al. J Clin Invest 102: 1125-1131, 1998
- Rawlins EL, et al. Proc Natl Acad Sci U S A 104: 410-417, 2007
- Borok Z, et al. Proc Am Thorac Soc 8: 215-222, 2011
- Hogan BL, et al. Cell Stem Cell 15: 123-138, 2014
- Kotton DN and Morrisey EE. Nat Med 20: 822-832, 2014
- Rock JR and Hogan BL. Annu Rev Cel Dev Biol 27: 493-512, 2011
- Rock JR, et al. Dis Model Mech 3: 545-556, 2010
- Stripp BR and Reynolds SD. Proc Am Thorac Soc 5: 328-333, 2008
- Wansleeben C, et al. Dev Biol 2: 131-148, 2013
- Hackett TL, et al. Stem Cells 26(10): 2576-85, 2008
- Reynolds SD, et al. Proc Am Thorac Soc 9: 27-37, 2012
- Rock JR, et al. Proc Natl Acad Sci U S A 106: 12771-12775, 2009
- Lee JH, et al. Cell 156: 440-455, 2014
- Barkauskas CE, et al. J Clin Invest 123: 3025-3036, 2013
- Cole BB, et al. Am J Pathol 177: 362-376, 2010
- Ghosh M, et al. Am J Respir Cell Mol Biol 45: 459-469, 2011
- Ghosh M, et al. Stem Cells 31: 2767-2778, 2013
- Ahmad S, et al. Am J Respir Cell Mol Biol 48: 94-104, 2013
- Ghosh M, et al. Am J Respir Cell Mol Biol 45: 403-410, 2011
- Ghosh M, et al. Am J Respir Cell Mol Biol 49: 1127-1134, 2013
- Tata PR, et al. Nature 503: 218-223, 2013
- Giangreco A, et al. Am J Pathol 161: 173-182, 2002
- Desai TJ, et al. Nature 507: 190-194, 2014
- Evans MJ, et al. Exp Mol Mathol 22: 142-150, 1975
- Evans MJ, et al. Am J Pathol 70: 175-198, 1973
- Kaplan HP, et al. Lab Invest 20: 94-100, 1969
- Berube K, et al. Toxicology 278: 311-318, 2010
- Muller L, et al. J Vis Exp 80, 2013
- Whitcutt MJ, et al. In Vitro Cell Dev Biol 24: 420-428, 1988
- Neuberger T, et al. Methods Mol Biol 741: 39-54, 2011
- Randell SH, et al. Methods Mol Biol 742: 285-310, 2011
- Pezzulo AA, et al. Am J Physiol Lung Cell Mol Physiol 300: L25-31, 2011
- Lin H, et al. J Pharm Sci 96: 341-350, 2007
- Lee MK, et al. Drug Deliv 12: 305-311, 2005
- Dvorak A, et al. Am J Respir Cell Mol Biol 44: 465-473, 2011
- Wong AP, et al. Nat Biotechnol 30(9): 876-82, 2012
- Kastner PE, et al. Toxicol In Vitro 27: 632-640, 2013
- Mathis C, et al. Am J Physiol Lung Cell Mol Physiol 304: L489- 503, 2013

