Monolayer Culture of Mouse Mammary Epithelial Cells
- Document # 29179
- Version 2.2.0
- Sep 2015
Mammary Epithelial Cell Hierarchy
The mammary gland is a compound, tubulo-alveolar gland composed of a series of branched ducts that drain sac-like alveoli (lobules) during lactation, surrounded by a fat pad in mice and by fibrous and fatty connective tissues in humans. Emerging evidence indicates that the mammary epithelium in both humans and mice may comprise a cellular hierarchy: from the mammary stem cell to various progenitors, and finally to differentiated luminal and myoepithelial cells. Two types of luminal cells line the mammary gland ducts and alveoli: those that express the estrogen receptor (ER), and those that do not (ER- ). Basal to the luminal cells and adjacent to the basement membrane are the myoepithelial cells: specialized contractile epithelial cells that express smooth muscle actin and smooth muscle myosin.
Electron microscopic studies demonstrate that approximately 3 - 8% of all mammary epithelial cells do not have a terminally differentiated phenotype, and are considered to reside higher within the mammary epithelial cell hierarchy.1,2 For instance, it is thought that ER- luminal cells in the virgin mammary gland are progenitor cells that proliferate during pregnancy to generate alveoli; those cells in the alveoli are then capable of producing milk during lactation.3 The most primitive cell in the hierarchy would be the mammary stem cell (MaSC), traditionally defined as a cell that can generate both the ductal and lobular structures of the mammary gland (complete with all the cell lineages), and can self-renew. However, as will be described below, evidence suggests that a new definition for the MaSC may be needed.
Mouse Mammary Stem Cells
In 1998, Kordon and Smith demonstrated that a single genetically marked mouse mammary epithelial cell could recapitulate both the ductal and lobular components of the mammary epithelium, as well as self-renew when transplanted into epithelium-free (cleared) mammary fat pads of primary and secondary recipient mice.4 Although such cells meet the functional criteria for MaSCs, they are operationally termed mammary repopulating units (MRUs). MRUs are known to be distributed throughout the mammary epithelium, since transplantation of any segment of the epithelium into surgically cleared (epitheliumfree) fat pads recapitulates the mammary epithelial tree, with a maximum of seven transplant generations before loss of repopulating ability.5-8 They occur with the highest frequency in the terminal end buds (the dilated end of the developing ducts during puberty), and with the lowest frequency in lactating alveoli.7 The phenotype of mouse MRUs was first correlated to in vivo repopulating ability by Welm and colleagues, who demonstrated that expression of Sca-1 enriches for MRUs.9 Subsequent studies demonstrated that it is the Sca-1low fraction of the Sca-1+ population that is enriched for MRUs, and that most MRUs have a basal (EpCAM+CD24+CD29hiCD49fhi) phenotype.10,11
Unexpectedly, MRUs express neither ER nor the progesterone receptor (PR). Both steroid hormones, particularly estrogen, are strong drivers of mammary gland development, and exposure to these hormones is associated with increased breast cancer risk.12 However, it is now emerging that estrogen and progesterone exert many of their effects in a paracrine fashion: by binding to their respective receptors in differentiated ER- and PR- expressing luminal cells, which then secrete factors such as RANK ligand, Wnt-4 and amphiregulin that act on the more primitive ERPR- cells of the mammary epithelium (e.g. MRUs).13-18 The correlation is reflected in the 14-fold fluctuation in MRU numbers during the mouse estrus cycle, with maximal MRU numbers during the luteal dioestrus phase when progesterone levels are highest.13 Steroid hormones such as progesterone may however also directly influence mammary epithelial cell growth by promoting proliferation of PR+ progenitor cells.19
This said, a recent study has challenged the equation of MRUs with MaSCs. In vivo lineage tracing experiments in mice have demonstrated that basal stem cells can generate all the cells of the mammary gland during prenatal development, but in the postnatal gland the luminal and basal (myoepithelial) lineages are maintained by lineage-specific stem cells.20 The precise identity and properties of these luminal stem cells in the mouse mammary gland is not currently known.
Electron microscopic studies have also identified small electron-lucent cells, known as small light cells or SLCs, as a potential candidate for MaSCs and their immediate primary progenitors.1,2 These cells are basally positioned within the mammary epithelium and occur at a frequency of 3%, but are absent from senescent epithelium.21 SLCs undergo mitotic division and give rise to cells of both luminal and myoepithelial phenotypes, suggesting they are not lymphocytes traversing the epithelium. But to date, no functional evidence of stem cell activity has yet been reported for SLCs, nor have strategies to genetically mark or isolate these cells been established. Another potential MaSC candidate are the cap cells lining the terminal end buds of the elongating mammary ducts during adolescence.22-24 These cells are present nowhere else, and they express both luminal and myoepithelial-related proteins at low levels. Some investigators have postulated that the cap cells represent the stem cells of the rodent mammary gland, although others have suggested that they are merely myoepithelial progenitors paving the way for ductal elongation during development.22-25 Like SLCs, the transitory cap cells have not been demonstrated in functional assays to have any ability to regenerate mammary epithelial structures.
Seeding of mammary epithelial cells into in vitro colony-forming unit (CFU) assays reveals that a large population of CFUs resides within the luminal cell compartment, and that most of these progenitor cells have a CD24hiSca-1-ER- phenotype.3,10,11 Transplantation of single-cell suspensions of mammary epithelial cells at limiting dilutions into cleared fat pads demonstrates the existence of ductal-restricted and lobular-restricted progenitors, both of which derive from the mammary stem cell.26,27 The frequency and phenotypic profile of these two structurally-restricted progenitors are not known. Another unique progenitor that has been described is the parity-induced epithelial progenitor that arises following the first pregnancy.27-29 These cells, which have previously undergone lactogenic differentiation, do not undergo apoptosis during mammary gland remodeling following pregnancy, but instead survive and are thought to act as self-renewing alveolar precursors in subsequent pregnancies.
1.0 Preparation of EpiCult™-B Medium (Mouse)
- Prepare complete EpiCult™-B Medium (Mouse) by adding 50 mL of thawed EpiCult™-B Proliferation Supplement (Mouse; Catalog #05612) to 450 mL of EpiCult™-B Basal Medium (Mouse; Catalog #05611).
Cytokines are not included in medium or supplements and must be added. STEMCELL Technologies recommends adding:- 10 ng/mL recombinant human Epidermal Growth Factor (rh EGF; Catalog #02633)
- 10 ng/mL recombinant human Basic Fibroblast Growth Factor (rh bFGF; Catalog #02634)
- 4 μg/mL (0.0004%) Heparin (Catalog #07980)
- Complete EpiCult™-B Medium (Mouse) without added cytokines is stable at 2 - 8°C for 1 month.
Cytokines should be added immediately before use. Avoid repeated exposure of medium to room temperature and light during experiments. If the entire volume is not needed immediately, aliquot into appropriate volumes to be used within 1 week (e.g. 100 mL).
This product does not contain antibiotics. If desired, add penicillin and streptomycin to 500 mL of medium to achieve a final concentration of 100 U/mL penicillin and 100 μg/mL of streptomycin. Following addition of antibiotics, medium should be used within 1 week.
2.0 Protocol for Culturing Mouse Mammary Epithelial Cells Using EpiCult™-B Medium (Mouse)
2.1 Dissociation of Mouse Mammary Tissue
- Prepare dissociation solution by diluting 1 part Collagenase/ Hyaluronidase (Catalog #07912) or Gentle Collagenase/ Hyaluronidase (Catalog #07919) with 9 parts Complete EpiCult™-B Medium (Mouse) supplemented with 5% FBS and place into a 15 mL or 50 mL centrifuge tube. Approximately 2 - 5 mL of the dissociation solution will be required for every 2 mammary glands to be dissociated.
- Resect mammary glands and transfer to a sterile glass petri dish. Mince with scalpels in a cross-wise pattern until glands are rendered to a paste. Transfer the mammary tissue to the tube containing the dissociation solution (prepared in Step 1) and incubate at 37°C for 6 - 8 hours (if using Collagenase/ Hyaluronidase) or overnight (if using Gentle Collagenase/ Hyaluronidase).
Important Note: Mouse mammary glands can be dissociated for shorter lengths of time (1 - 2 hours), however the mammary stem cell yield is dramatically (~80%) reduced (and in some cases, absent). A short tissue dissociation time results in a high yield of liberated lymphocytes and stromal cells, but the stem cells are apparently lost in the filtration stage (Section 2.2, Step 6), even following trypsin and dispase treatment.
Conversely, digestion longer than 8 hours in Collagenase/ Hyaluronidase results in over-digestion of the cells and a decrease in stem cell yield (but not total cell yield). With Gentle Collagenase/Hyaluronidase, the mouse mammary tissue can be dissociated overnight (for up to 15 hours) without compromising mouse mammary stem and progenitor cell content. - After dissociation, centrifuge the cells at 350 x g for 5 minutes and discard the supernatant.
- Resuspend the pellet with a 1:4 mixture of cold Hanks’ Balanced Salt Solution Modified (Catalog #37150) supplemented with 2% FBS and Ammonium Chloride Solution (Catalog #07800) and centrifuge at 350 x g for 5 minutes. The resultant pellet contains epithelial cell organoids as well as stromal cells and lymphocytes. To generate a single-cell suspension of mammary epithelial cells, refer to Section 2.2.
2.2 Generation of Single-Cell Suspensions From Dissociated Mouse Mammary Tissue
- Add 1 - 5 mL of pre-warmed Trypsin-EDTA (Catalog #07901) to the partially-dissociated tissue (generated in Section 2.1) and mix by pipetting.
- Gently pipette up and down with a P1000 micropipettor for 1 - 3 minutes. The sample should become very stringy due to lysis of dead cells and the release of DNA.
- Add 10 mL of cold Hanks’ Balanced Salt Solution Modified (Catalog #37150) supplemented with 2% FBS and centrifuge at 350 x g for 5 minutes. The Hanks’ + FBS solution is now referred to as HF.
- Remove as much of the supernatant as possible.
- Add 2 mL of pre-warmed Dispase (5 U/mL; Catalog #07913) and 200 µL of DNase I Solution (1 mg/mL; Catalog #07900). Pipette the sample for 1 minute with a P1000 micropipettor to further dissociate cell clumps. The sample should now be cloudy, but not stringy. If still stringy, add an additional 100 µL of DNase I Solution.
- Dilute the cell suspension with an additional 10 mL of cold HF and filter the cell suspension through a 40 µm Cell Strainer (Catalog #27305) into a new 50 mL centrifuge tube. Centrifuge 350 x g for 5 minutes and discard the supernatant.
- If the cell pellet is heavily contaminated with red blood cells, resuspend the pellet in a 1:4 mixture of cold HF:Ammonium Chloride Solution (Catalog #07800), centrifuge at 350 x g for 5 minutes and discard the supernatant.
2.3 Culture of Mouse Mammary Epithelial Cells
Mouse mammary epithelial cell cultures should be initiated from single cell suspensions (refer to Section 2.2). Otherwise, cells will not adhere well to the tissue culture flask.
- Seed mouse mammary cells into tissue culture flasks at a density of 2 - 4 x 103 cells/cm2 in Complete EpiCult™-B Medium (Mouse) supplemented with rh EGF (Catalog #02633), rh bFGF (Catalog #02634) and Heparin (Catalog #07980) (as described in Section 1.0) and 5% FBS (Catalog #06100). For example, in a T-75 cm2 flask (Corning; Catalog #03276) use 1.5 - 3.0 x 105 cells. For a 6-well plate (VWR; Catalog #62406-161), use 2 - 4 x 104 cells/well.
Failure to include serum during plating of mouse mammary epithelial progenitor cells will result in poor adherence of the cells to the tissue culture plastic. - After 24 hours, change the culture medium to serum-free Complete EpiCult™-B Medium (Mouse) containing cytokines.
Failure to change the medium to serum-free Complete EpiCult™-B Medium (Mouse) could result in overgrowth of the culture by contaminating stromal cells.
Mammary epithelial cells can be subcultured by first washing the adherent cells with Hanks’ Balanced Salt Solution Modified (Catalog #37150) followed by incubation with pre-warmed Trypsin-EDTA (Catalog #07901). Once the cells have detached from the culture vessel, an equal volume of cold HF (refer to Section 2.2, Step 3) should be added and the cell suspension centrifuged at 350 x g. Collected cells can then be reseeded into tissue culture flasks as described above in Steps 1 and 2.
It is recommended that the cultures do not exceed 80% confluence. While some biological variability does exist, under described conditions the mammary culture will typically be ready for subculture 7 - 10 days after seeding.
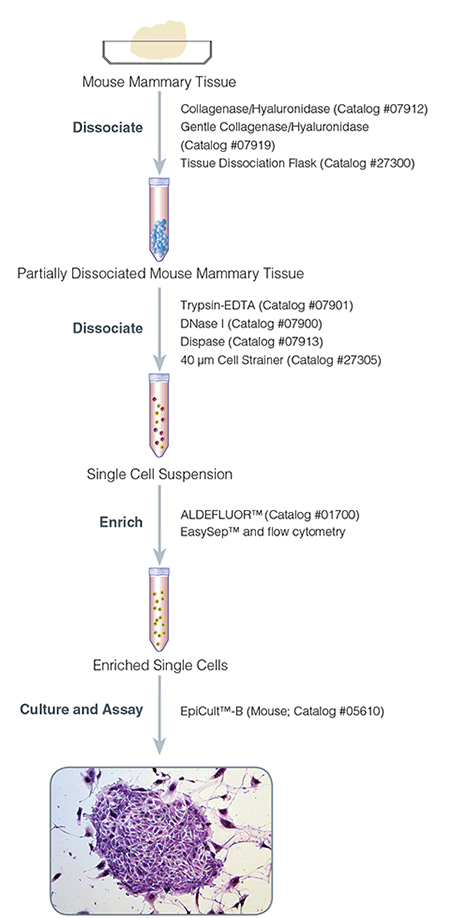
Figure 1. Protocol for Isolation and Culture of Mouse Mammary Epithelial Cells
Phase contrast photographs of a prototypical mouse colony. Unlike human mammary CFC colonies, mouse mammary epithelial cell colony subtypes are not easily identifiable. The colonies were visualized by Wright-Giemsa staining. Mouse colonies were obtained in EpiCult™-B (Mouse) and cultured in the presence of an irradiated NIH 3T3 feeder layer.
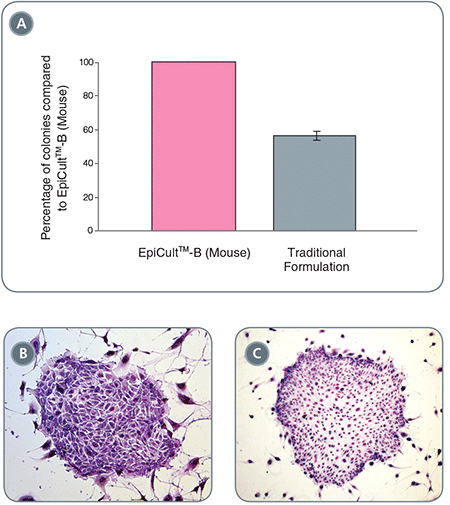
Figure 2. EpiCult™-B Mouse Mammary Epithelial Cell Colonies
(A) EpiCult™-B (Mouse) generates twice as many colonies compared to commonly used formulation. (B) and (C) are examples of mouse mammary epithelial colonies grown in EpiCult™-B (Mouse).
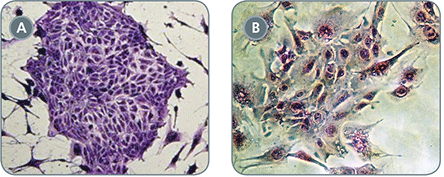
Figure 3. EpiCult™-B (Mouse) Cell Cultures Display Decreased EMT
Mammary epithelial cells display a clear delay and decrease of epithelial-to-mesenchymal transition (EMT) when cultured in (A) EpiCult™-B (Mouse) versus (B) traditional formulation (photo courtesy of Dr. John Stingl). EMT is defined by emergence of a fibroblastic-like morphology.
Detect Mouse Mammary Colony-Forming Units

RECOMMENDED FOR:
Colony-forming unit assays with mouse mammary epithelial cells. Promotes the clonogenic growth of mouse mammary progenitors when used in conjuction with an irradiated feeder layer (NIH 3T3). Also used for the enzymatic dissociation of mouse mammary tissue when supplemented with Collagenase/Hyaluronidase (Catalog #07912) or Gentle Collagenase/Hyaluronidase (Catalog #07919).
CONTAINS:
- EpiCult™-B Basal Medium (Mouse): 450 mL
- EpiCult™-B Proliferation Supplements (Mouse): 50 mL
REQUIRES:
Supplement with 10 ng/mL rh Epidermal Growth Factor (Catalog #02633), 10 ng/mL rh Basic Fibroblast Growth Factor (Catalog #02634) and 4 μg/mL Heparin (Catalog #07980) before use.
References
- Chepko G, et al. Tissue Cell 29: 239-253, 1997
- Chepko G, et al. J Mammary Gland Biol Neoplasia 4: 35-52, 1999
- Sleeman KE, et al. J Cell Biol 176:19-26, 2007
- Kordon EC, et al. Development 125: 1921-1930, 1998
- Hoshino K. Anat Rec 151: 364, 1965
- Daniel CW, et al. Proc Natl Acad Sci USA 61: 53-60, 1968
- Smith GH, et al. J Cell Sci 90: 173-183, 1988
- Daniel CW, et al. Exp Cell Res 65: 27-32, 1971
- Welm BE, et al. Dev Biol 245: 42-56, 2002
- Stingl J, et al. Nature 439: 993-997, 2006
- Shackleton M, et al. Nature 439: 84-88, 2006
- Asselin-Labat M-L, et al. J Natl Cancer Inst 98: 1011-1014, 2006
- Joshi, PA, et al. Nature 465: 803-807, 2010
- Asselin-Labat M-L, et al. Nat Cell Biol 9:201-209, 2007
- Wilson CL, et al. Endocr-Relat Cancer 13: 617-628, 2006
- Mallepell S, et al. Proc Natl Acad Sci U S A 103: 2196-2201, 2006
- Booth BW, et al. Exp Cell Res 316: 422-432, 2010
- Brisken C, et al. Genes Dev 14: 650-654, 2000
- Beleut M, et al. Proc Natl Acad Sci U S A 107: 2989-2994, 2010
- Van Keymeulen A, et al. Nature 479: 189-193, 2011
- Smith GH, et al. Cell Tissue Res 310: 313-320, 2002
- Williams JM, et al. Dev Biol 97: 274-290, 1983
- Rudland PS. J Histochem Cytochem 39: 1471-1484, 1991
- Rudland PS. Histol Histopathol 8: 385-404, 1993
- Sapino A, et al. Differentiation 55: 13-18, 1993
- Smith GH. Breast Cancer Res Treat 39: 21-31, 1996
- Smith GH, et al. Cell Prolif 36 supplement: 3-15, 2003
- Wagner K-U, et al. Development 129: 1377-1386, 2002
- Boulanger CA, et al. Oncogene 24: 552-560, 2005
Request Pricing
Thank you for your interest in this product. Please provide us with your contact information and your local representative will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration

