Hepatic Organoids

Hepatic Organoids
Hepatic organoids are functional three-dimensional (3D) in vitro models of the liver and serve as a novel platform to address diverse research questions pertinent to hepatic development and regeneration, detoxification and metabolism studies, liver disease modelling, and adult stem cell biology. Hepatic organoids consist of a spherical monolayer epithelium, preserve key physiological features of the liver and are obtained through isolation and expansion of stem and progenitor cells from hepatic stem cell niches.
By presenting an in vitro system that closely mimics an in vivo environment, hepatic organoids are enabling scientists to investigate their specific research questions in a physiologically relevant context.
The Liver
Hepatic Homeostasis, Regeneration and Response to Injury
While the stem cell niche in epithelial organs such as the intestine is highly defined, there is an ongoing debate in the liver research field about the source and contribution of stem cells, progenitor cells and mature cell types in the context of liver homeostasis and regeneration upon injury.5–7 Proliferative cells, localized in the periportal and pericentral regions, respond to different forms of liver injury (i.e. acute versus chronic).8 Besides the differentiation of “classical" stem cells or progenitors within the hepatic lineage to functional liver cells, it has also been postulated that existing, mature parenchymal cells contribute to tissue turnover.9
In tissue homeostasis, the liver has a slower turnover rate than other epithelial organs; the life-expectancy of a mature hepatocyte has been dated to 201 days.10 Homeostatic self-renewal and maintenance of hepatic function has recently been attributed to proliferative Axin2+ cells found in close proximity to the central vein within the liver lobule.11 Expression of Tbx3 in this cell type suggests the retention of an embryonic hepatocyte progenitor and demonstrates the presence of a heterogeneous and dynamic population of mature hepatocytes and hepatic progenitors within the adult liver. Fueled by Wnt signaling factors from neighbouring endothelial cells, these hepatic progenitors are maintained within the pericentral niche and give rise to progeny hepatocytes (but not cholangiocytes) that can reconstitute 40% of hepatocytes within the hepatic lobule.11 Other studies have localized a self-renewing hepatocyte progenitor population to the periportal region of the hepatic lobe, suggesting a secondary source for hepatic regeneration under normal conditions.12,13 These cells are proposed to gradually stream from the portal space toward the hepatic vein while undergoing different states of maturity and function during their journey. Additionally, Furuyama and colleagues have reported self-renewing Sox9-expressing progenitors in the biliary ductal epithelium as the principal source of cell replacement.14 Sox9 cell-derived hepatocytes were found to also travel from the periportal space to the central vein. Furthermore, derivatives of the Sox9 progenitors were localized to the extrahepatic bile duct tree, suggesting bipotent behavior of these progenitors. In contrast, others were able to attribute solely mature hepatocytes to liver homeostasis.15
The liver is well-adapted to respond to specific injury stimuli with fine-tuned responses of regeneration. As such, the surgical removal of a majority of the liver (partial hepatectomy) results in restoration of hepatic mass and function within a week.16 This suggests mitotic behavior of all cell populations in the periportal zone, including hepatocytes and bile duct cells, and this proliferation is believed to be activated in response to hepatic mitogens such as NFκB, STAT3, Jun and Fos. The responsiveness of hepatocytes and bile duct cells to readily divide in specific circumstances has dispelled the myth that hepatic parenchymal cells are terminally differentiated. However, in fibrotic and cirrhotic injury responses that impair the proliferative capacity of mature liver cell types, facultative stem cells of the liver may be activated to generate intermediate, duct-like “oval cells" that can subsequently restore function and liver mass.17 Lineage-tracing experiments have mapped this regeneration potential to the canal of Hering in the periportal regions of hepatic lobules.18 While coined “oval cell proliferation" in rodent models, this response is known as “ductular reaction" in human.19 Furthermore, progenitors expressing the transcription factor Sox9 are not only involved in tissue homeostasis, but have also been shown to contribute to the restoration of hepatic mass and function upon partial hepatectomy and acute liver injury (e.g. via CCl4 administration).14
Derivation of Liver Organoids from Adult Liver Stem Cells
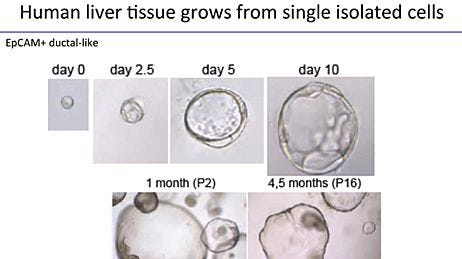
While Lgr5 stem cell marker-expression is generally not detected in the adult wild type liver, injury triggers the activation of an epithelial Lgr5+ epithelial progenitor cell population.20 Isolation of single Lgr5+ cells from the adult mouse liver and their growth into hepatic organoids has demonstrated their clonogenic potential and the long-term culture of these organoids has confirmed their indefinite capacity for self-renewal.20 Cells maintain their differentiation potential and exhibit karyotype stability. For human cultures, Epcam+ bile duct-derived bipotent progenitors were isolated for expansion into human hepatic organoids.21
Liver organoids differ from intestinal organoids in their limitation to undergo spontaneous differentiation during maintenance and expansion. Protocols for the propagation of liver organoids therefore rely on culture conditions for the establishment and expansion of hepatic organoids, followed by a second culture environment that removes proliferative signals, inhibits ductal lineage commitment and induces differentiation towards hepatocyte-like cells.20 Current differentiation protocols yield organoids of which 30-50% fulfill hepatic characteristics.22 These hepatic organoids resemble primary hepatocytes but lack the complete functional repertoire of this target cell type, a drawback shared by the hPSC-to-hepatocyte differentiation field. During expansion conditions, hepatic organoids express a ductal phenotype, characterized by the expression of Krt19. However, upon differentiation, hepatic organoids downregulate ductal markers and exhibit albumin secretion, CYP3A activity, LDL uptake and glycogen storage. Transplantation of differentiated hepatic organoids into mouse models of liver failure have shown engraftment and repopulation of the damaged liver, resulting in partial rescue of the murine liver function.20 Equivalent human liver organoids engrafted into mice with acute liver damage resulted in human albumin and alpha-1-antitrypin synthesis that resembled levels of secretions seen from transplants using primary human hepatocytes within one month.21 Furthermore, the disease pathogenesis of alpha-1-antitrypsin deficiency and Alagille syndrome were confirmed in human liver organoids derived from patient biopsies, showing proof-of-principle that diseased tissue can be cultured in 3D ex vivo, differentiated and utilized for disease modeling.21
The establishment of hepatic organoids from different species requires specialised and optimised media formulations (e.g. mouse, rat, human, dog).23,24
Derivation of Liver Organoids from Human Pluripotent Stem Cells
3D liver tissue has also been engineered by using using human induced pluripotent stem cells (hiPSCs) to derive hepatocytes in co-culture with mesenchymal and endothelial cells.25 Mimicking organogenesis, cells self-organized into liver buds and when transplanted into mice, liver buds were vascularized and matured to synthesize serum proteins and carry out detoxifying functions. In a mouse model of liver failure, transplanted liver buds were able to restore partial hepatic function and rescue affected mice. In efforts toward clinical applications, current research aims to design liver buds suitable for hepatic delivery via the portal vein of patients in need of a liver transplant.26
Protocols have also been established to differentiation hPSCs to cholangiocyte-like cells.27,28 Using 3D culture conditions, bile duct cells polarized into cystic organoids and exhibited a morphology and functionality reminiscent of primary bile duct cells. These culture systems have shown suitability to model cholangiocyte-specific conditions such as Alagille syndrome, polycystic liver disease and cystic fibrosis-associated cholangiopathies.27 Furthermore, cholangiocyte-like cells in 3D environments were used to validate novel therapeutic agents and identify drug targets that confine the associated pathophysiologies.27
One drawback of hIPSCs or trans-differentiated cells in the design of clinical solutions is their exposure to genetic modifications through reprogramming factors. While these cells commonly exhibit genomic instability, particularly in long-term cultures, adult stem cells directly derived from hepatic tissue circumvent this caveat.29
Applications for Liver Organoids
An improved understanding of hepatic regenerative pathways and the development of in vitro systems to mimic both the expansion and differentiation of hepatocytes, will pave the way towards autologous cell therapy and transplantation.30 The derivation of hepatic organoids from patients with inherited liver disease also opens the opportunity to establish drug-screening platforms and toxicity assays for personalized medicine while allowing researchers to further dissect the signaling pathways involved in the underlying disease.31 Genome editing technologies coupled with the liver organoid culture platform also allows the reintroduction of genetically corrected stem cells into affected patients where they may differentiate to healthy hepatocytes capable of their wild type functionalities.32
- Desmet V. (1994) Organizational Principles. In: Arias IM et al. (Eds), The liver: biology and pathobiology. (pp 3-14). Philadelphia, PA: Lippencott-Raven Publishers.
- Lemaigre F & Zaret KS. (2004) Liver development update: New embryo models, cell lineage control, and morphogenesis. Curr Opin Genet Dev 14(5): 582–90.
- Wisse E et al. (1996) Structure and function of sinusoidal lining cells in the liver. Toxicol Pathol 24(1): 100–11.
- Kanno N et al. (2000) Functional heterogeneity of the intrahepatic biliary epithelium. Hepatology: 555–61.
- Sell S. (1990) Is there a liver stem cell? Cancer Res (50): 3811–5.
- Thorgeirsson SS. (1996) Hepatic stem cells in liver regeneration. Faseb J 10(11): 1249–56.
- Kopp JL et al. (2016) Stem cells versus plasticity in liver and pancreas regeneration. Nat Cell Biol 18(3): 238–45.
- Duncan A et al. (2009) Stem cells and liver regeneration. Gastroenterology 137(2): 466–81.
- Ponder K. (1996) Analysis of liver carcinogenesis by genetic marking studies. FASEB J 10(7): 673–82.
- Zajicek G et al. (1985) The streaming liver. Liver 5(6): 293–300.
- Wang B et al. (2015) Self-renewing diploid Axin2+ cells fuel homeostatic renewal of the liver. Nature 524(7564): 180–5.
- Fellous TG et al. (2009) Locating the stem cell niche and tracing hepatocyte lineages in human liver. Hepatology 49(5): 1655–63.
- Steiner JW et al. (1966) Cell population dynamics in the liver. A review of quantitative morphological techniques applied to the study of physiological and pathological growth. Exp Mol Pathol 5(2): 146–81.
- Furuyama K et al. (2011) Continuous cell supply from a Sox9-expressing progenitor zone in adult liver, exocrine pancreas and intestine. Nat Genet 43(1): 34–41.
- Español-Suñer R et al. (2012) Liver progenitor cells yield functional hepatocytes in response to chronic liver injury in mice. Gastroenterology 143(6): 1564–75.
- Taub R. (2004) Liver regeneration: from myth to mechanism. Nat Rev Mol Cell Biol 5(10): 836–47.
- Alison MR et al. (2004) Hepatic stem cells: From inside and outside the liver? Cell Prolif 37(1): 1–21.
- Theise ND et al. (1999) The canals of Hering and hepatic stem cells in humans. Hepatology 30(6): 1425–33.
- Popper H et al. (1957) Ductular cell reaction in the liver in hepatic injury. J Mt Sinai Hosp N Y 24(5): 551–6.
- Huch M et al. (2013) In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature 494(7436): 247–50.
- Huch M et al. (2015) Long-term culture of genome-stable bipotent stem cells from adult human liver. Cell 160(1–2): 299–312.
- Huch M et al. (2013) Lgr5(+) liver stem cells, hepatic organoids and regenerative medicine. Regen Med 8(4): 385–7.
- Kuijk EW et al. (2016) Generation and characterization of rat liver stem cell lines and their engraftment in a rat model of liver failure. Sci Rep 6: 22154.
- Nantasanti S et al. (2015) Disease modeling and gene therapy of copper storage disease in canine hepatic organoids. Stem Cell Reports 5(5): 895–907.
- Takebe T et al. (2013) Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 499(7459): 481–4.
- Willyard C. (2015)The boom in mini stomachs, brains, breasts, kidneys and more.. Nature 523(7562): 520–2.
- Sampaziotis F et al. (2015) Cholangiocytes derived from human induced pluripotent stem cells for disease modeling and drug validation. Nat Biotechnol 33(8): 845–52.
- Ogawa M et al. (2015) Directed differentiation of cholangiocytes from human pluripotent stem cells. Nat Biotechnol 33(8): 853–61.
- Huch M & Koo B-K. (2015) Modeling mouse and human development using organoid cultures. Development 142(18): 3113–25.
- Willenbring H & Soto-Gutierrez A. (2013) Transplantable liver organoids made from only three ingredients. Cell Stem Cell 13(2): 139-40.
- Au SH et al. (2014) Hepatic organoids for microfluidic drug screening. Lab Chip 14(17): 3290–9.
- Fatehullah A et al. (2016) Organoids as an in vitro model of human development and disease. Nat Cell Biol 18(3): 246–54.

Researcher Profile: Meritxell Huch, PhD
Principal Investigator at the Wellcome Trust/CRUK Gurdon Institute

Webinar: From Stem Cells to Organoids
Dr. Meritxell Huch discusses the use of organoids to model the GI tract
Key Applications of Hepatic Organoids
Primary Cell/Tissue-derived Liver Organoids and other Three Dimensional Liver Structures
Huch M, Gehart H, van Boxtel R, Hamer K, Blokzijl F, Verstegen MMA, Ellis E, van Wenum M, Fuchs SA, de Ligt J et al. (2015) Long-Term Culture of Genome-Stable Bipotent Stem Cells from Adult Human Liver. Cell 160(1–2): 299–312.
Huch M et al. (2013) In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature 494(7436): 247–50.
Kasuya J, Sudo R, Masuda G, Mitaka T, Ikeda M & Tanishita K. (2015) Reconstruction of hepatic stellate cell-incorporated liver capillary structures in small hepatocyte tri-culture using microporous membranes. J Tissue Eng Regen Med 9(3): 247–256.
Kuijk EW, Rasmussen S, Blokzijl F, Huch M, Gehart H, Toonen P, Begthel H, Clevers H, Geurts AM & Cuppen E. (2016) Generation and characterization of rat liver stem cell lines and their engraftment in a rat model of liver failure. Sci Rep 6: 22154
Nantasanti S et al. (2015) Disease modeling and gene therapy of copper storage disease in canine hepatic organoids. Stem Cell Reports 5(5): 895–907.
Ramachandran SD, Schirmer K, Münst B, Heinz S, Ghafoory S, Wölfl S, Simon-Keller K, Marx A, Øie CI, Ebert MP et al. (2015) In Vitro Generation of Functional Liver Organoid-Like Structures Using Adult Human Cells. Avila MA (Ed). PLoS One 10(10)
Takebe T, Sekine K, Suzuki Y, Enomura M, Tanaka S, Ueno Y, Zheng Y-W & Taniguchi H. (2012) Self-Organization of Human Hepatic Organoid by Recapitulating Organogenesis In Vitro. Transplant Proc 44(4): 1018–1020.
hPSC-derived Liver Organoids
Guye P, Ebrahimkhani MR, Kipniss N, Velazquez JJ, Schoenfeld E, Kiani S, Griffith LG & Weiss R. (2016) Genetically engineering self-organization of human pluripotent stem cells into a liver bud-like tissue using Gata6. Nat Commun 7: 10243
Takebe T, Sekine K, Enomura M, Koike H, Kimura M, Ogaeri T, Zhang RR, Ueno Y, Zheng YW, Koike N, Aoyama S, Adachi Y, Taniguchi H. (2013) Vascularized and functional human liver from an iPSC-derived organ bud transplant.. Nature. 499(7459:481-4))
Takebe T, Zhang R-R, Koike H, Kimura M, Yoshizawa E, Enomura M, Koike N, Sekine K & Taniguchi H. (2014) Generation of a vascularized and functional human liver from an iPSC-derived organ bud transplant. Nat Protoc 9(2): 396–409.
Ichinohe N, Tanimizu N, Ooe H, Nakamura Y, Mizuguchi T, Kon J, Hirata K & Mitaka T. (2013) Differentiation capacity of hepatic stem/progenitor cells isolated from D -galactosamine-treated rat livers. Hepatology 57(3): 1192–1202.
Oval Cell-derived Hepatocyte Organoids
Liver Stem Cell Niche
Kopp JL et al. (2016) Stem cells versus plasticity in liver and pancreas regeneration. Nat Cell Biol 18(3): 238–245.
Miyajima A et al. (2014) Stem/Progenitor Cells in Liver Development, Homeostasis, Regeneration, and Reprogramming. Cell Stem Cell 14(5): 561–574.
Tarlow BD, Finegold MJ & Grompe M. (2014) Clonal tracing of Sox9 + liver progenitors in mouse oval cell injury. Hepatology 60(1): 278–289.
Yimlamai D, Christodoulou C, Galli GG, Yanger K, Pepe-Mooney B, Gurung B, Shrestha K, Cahan P, Stanger BZ & Camargo FD. (2014) Hippo Pathway Activity Influences Liver Cell Fate. Cell 157(6): 1324–1338.
Wang B et al. (2015) Self-renewing diploid Axin2+ cells fuel homeostatic renewal of the liver. Nature 524(7564): 180–185.
Microfluidic Organ-on-a-chip Technology to Model Liver Function
Gröger M, Rennert K, Giszas B, Weiß E, Dinger J, Funke H, Kiehntopf M, Peters FT, Lupp A, Bauer M et al. (2016) Monocyte-induced recovery of inflammation-associated hepatocellular dysfunction in a biochip-based human liver model. Sci Rep 6: 21868
Kobayashi A, Yamakoshi K, Yajima Y, Utoh R, Yamada M & Seki M. (2013) Preparation of stripe-patterned heterogeneous hydrogel sheets using microfluidic devices for high-density coculture of hepatocytes and fibroblasts. J Biosci Bioeng 116(6): 761–767.
Rennert K, Steinborn S, Gröger M, Ungerböck B, Jank A-M, Ehgartner J, Nietzsche S, Dinger J, Kiehntopf M, Funke H et al. (2015) A microfluidically perfused three dimensional human liver model. Biomaterials 71: 119–131
Schepers A, Li C, Chhabra A, Seney BT & Bhatia S. (2016) Engineering a perfusable 3D human liver platform from iPS cells. Lab Chip 16(14): 2644–53.
Liver Organoids as Models in Health and Disease
Leite SB, Roosens T, El Taghdouini A, Mannaerts I, Smout AJ, Najimi M, Sokal E, Noor F, Chesne C & van Grunsven LA. (2016) Novel human hepatic organoid model enables testing of drug-induced liver fibrosis in vitro. Biomaterials 78: 1–10
Lu Y, Zhang G, Shen C, Uygun K, Yarmush ML & Meng Q. (2012) A novel 3D liver organoid system for elucidation of hepatic glucose metabolism. Biotechnol Bioeng 109(2): 595–604.
Mavila N, Trecartin A, Spurrier R, Xiao Y, Hou X, James D, Fu X, Truong B, Wang C, Lipshutz GS et al. (2016) Functional Human and Murine Tissue-Engineered Liver Is Generated From Adult Stem/Progenitor Cells. Stem Cells Transl Med
Nantasanti S, Spee B, Kruitwagen HS, Chen C, Geijsen N, Oosterhoff LA, van Wolferen ME, Pelaez N, Fieten H, Wubbolts RW et al. (2015) Disease Modeling and Gene Therapy of Copper Storage Disease in Canine Hepatic Organoids. Stem Cell Reports 5(5): 895–907.
Sampaziotis F et al. (2015) Cholangiocytes derived from human induced pluripotent stem cells for disease modeling and drug validation. Nat Biotechnol 33(8): 845–852.
Skardal A, Devarasetty M, Rodman C, Atala A & Soker S. (2015) Liver-Tumor Hybrid Organoids for Modeling Tumor Growth and Drug Response In Vitro. Ann Biomed Eng 43(10): 2361–2373.
Soto-Gutierrez A, Navarro-Alvarez N, Yagi H, Nahmias Y, Yarmush ML & Kobayashi N. (2010) Engineering of an Hepatic Organoid to Develop Liver Assist Devices. Cell Transplant 19(6): 815–822.
Struecker B et al. (2015) Porcine Liver Decellularization Under Oscillating Pressure Conditions: A Technical Refinement to Improve the Homogeneity of the Decellularization Process. Tissue Eng Part C Methods 21(3): 303–313.
Takai A, Fako V, Dang H, Forgues M, Yu Z, Budhu A & Wang XW. (2016) Three-dimensional Organotypic Culture Models of Human Hepatocellular Carcinoma. Sci Rep 6: 21174
Review Papers
Clevers H. (2016)Modeling Development and Disease with Organoids.16;165(7):1586-97
Handa K, Matsubara K, Fukumitsu K, Guzman-Lepe J, Watson A & Soto-Gutierrez A. (2014) Assembly of Human Organs from Stem Cells to Study Liver Disease. Am J Pathol 184(2): 348–357.
Hindley CJ, Cordero-Espinoza L & Huch M. (2016) Organoids from adult liver and pancreas: Stem cell biology and biomedical utility. Dev Biol 420(2): 251–261.
Huch M, Boj SF & Clevers H. (2013) Lgr5 + liver stem cells, hepatic organoids and regenerative medicine. Regen Med 8(4): 385–387.
Nantasanti S, de Bruin A, Rothuizen J, Penning LC, Schotanus BA. (2016)Concise Review: Organoids Are a Powerful Tool for the Study of Liver Disease and Personalized Treatment Design in Humans and Animals. Stem Cells Transl Med. 5(3):325-30
Rashid T, Takebe T & Nakauchi H. (2015) Novel strategies for liver therapy using stem cells. Gut 64(1): 1–4.
Vacanti JP & Kulig KM. (2014) Liver cell therapy and tissue engineering for transplantation. Semin Pediatr Surg 23(3): 150–155.